 Mecanismos de transporte
Mecanismos de transporte
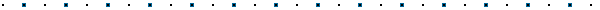
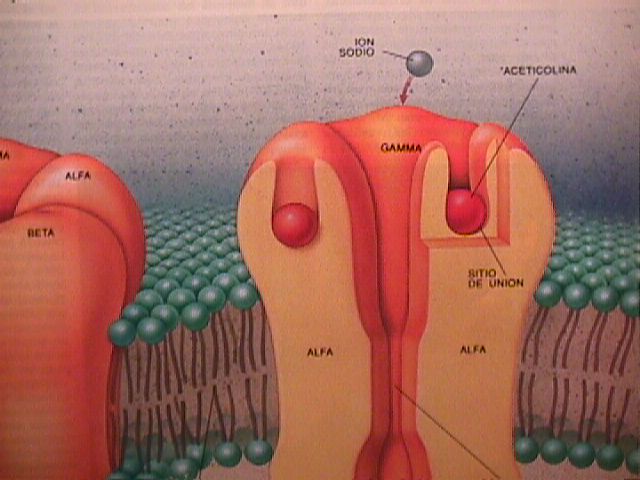
Es de capital importancia para la célula poder transportar
moléculas hacia afuera y adentro de ella misma. Imagine una proteína que tiene múltiples dominios transmembrana
(la atraviesan) y dispone los mismos en circulo formando un
cilindro o mejor un barril, que visto desde afuera, muestra cada
uno de los dominios, equivalentes a un listón del barril.
El "centro" de este barril conforma un "agujero" en la membrana
plasmática, aislado de la misma por un arreglo de dominios de
transmembrana alrededor de él. Este "agujero" puede ser utilizado
para transportar substancias hacia adentro o afuera de la
célula.
Este agujero puede ser hidrofílico si cadenas
laterales hidrofílicas de las proteínas que lo rodean protruyen
hacia él.
En la practica, para una proteína de membrana de estructura
conocida, estos agujeros solo son lo suficientemente grandes para
dejar pasar por la membrana plasmática moléculas pequeñas
tales como H+, K+ o Na+.
Estos iones pueden pasar por el orificio por difusión pasiva,
en cuyo caso la proteína que permite el paso conforma un
"canal iónico". En otros casos la proteína de
membrana necesita invertir energía (generalmente derivada de
ATP), para forzar el paso del ión de un lado al otro de la
membrana, en ese caso conforma una "bomba de iones".
Dado la importancia del transporte a través de la membrana la
célula utiliza un gran numero de mecanismos de transporte. Estos
mecanismos caen dentro de una de estas tres categorías:
difusión simple, difusión facilitada, y transporte activo.
Difusión simple, significa que la molécula puede pasar
directamente a través de la membrana. La difusión es siempre a
favor de un gradiente de concentración. Esto limita la
máxima concentración posible en el interior de la célula (o en
el exterior si se trata de un producto de desecho).
La efectividad de la difusión está limitada por la velocidad
de difusión de la molécula.
Por lo tanto si bien la difusión es un mecanismo de
transporte suficientemente efectivo para alguna moléculas (por
ejemplo el agua), la célula debe utilizar otros mecanismo de
transporte para sus necesidades.
La difusión facilitada utiliza canales (formados por
proteínas de membrana) para permitir que moléculas cargadas
(que de otra manera no podrían atravesar la membrana) difundan
libremente hacia afuera y adentro de la célula. Estos canales
son usados sobre todo por iones pequeños tales como K+, Na+,
Cl-.
La velocidad del transporte facilitado esta limitado por el
numero de canales disponibles (ver que la curva indica una
"saturación") mientras que la velocidad de difusión
depende solo del gradiente de concentración.
El transporte activo requiere un gasto de energía para
transportar la molécula de un lado al otro de la membrana, pero
el transporte activo es el único que puede transportar
moléculas contra un gradiente de concentración, al igual
que la difusión facilitada el transporte activo esta limitado
por el numero de proteínas transportadoras presentes.
Son de interés dos grandes categorías de transporte activo,
primario y secundario. El transporte activo primario usa energía
(generalmente obtenida de la hidrólisis de ATP), a nivel de la
misma proteína de membrana produciendo un cambio conformacional
que resulta en el transporte de una molécula a través de la
proteína.
El ejemplo mas conocido es la bomba de Na+/K+. La bomba de
Na+/K+ realiza un contratransporte("antyport")
transporta K+ al interior de la célula y Na+ al exterior de la
misma, al mismo tiempo, gastando en el proceso ATP.
El transporte activo secundario utiliza la energía
para establecer un gradiente a través de la membrana celular, y
luego utiliza ese gradiente para transportar una molécula de
interés contra su gradiente de concentración.
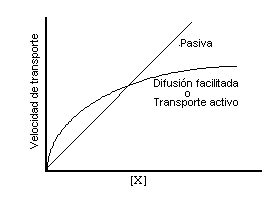
Un ejemplo de ese mecanismo es el siguiente: Escherichia
coli establece un gradiente de protones (H+) entre ambos
lados de la membrana utilizando energía para bombear protones
hacia afuera de la célula. Luego estos protones se acoplan a la
lactosa (un azúcar que sirve de nutriente al microorganismo) a
nivel de la lactosa-permeasa (otra proteína de transmembrana),
la lactosa permeasa usa la energía del protón moviéndose a
favor de su gradiente de concentración para transportar la
lactosa dentro de la célula.
Este transporte acoplado en la misma dirección a través de
la membrana celular se denomina cotransporte
("symport"). Escherichia coli utiliza este
tipo de mecanismo para transportar otros azucares tales como
ribosa y arabinosa, como así también numerosos aminoácidos.
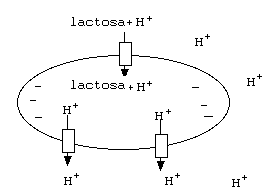
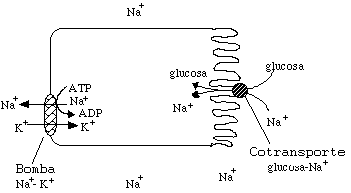
Otro sistema de transporte secundario usa la bomba de
Sodio/Potasio en una primera etapa, genera así un fuerte
gradiente de Sodio a través de la membrana. Luego la proteína
"simport" para el sistema Sodio-Glucosa usa la energía
del gradiente de Sodio para transportar Glucosa al interior de la
célula.
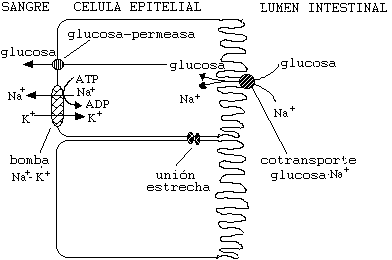
Este sistema se usa de manera original en las células
epiteliales del intestino. Estas células toman glucosa y sodio
del intestino y lo transportan al torrente sanguíneo utilizando
la acción concertada de los "simport" para
Sodio/Glucosa, la glucosa permeasa ( una proteína de difusión
facilitada para la glucosa) y las bombas de Sodio/Potasio.
Se debe hacer notar que las células del intestino se
encuentran unidas entre sí por "uniones estrechas" (tight junctions) que impiden que nada proveniente del intestino
pase al torrente sanguíneo sin ser primero filtradas por las
células epiteliales.
Algunas sustancias más grandes como polisacáridos,
proteínas y otras células cruzan las membranas plasmáticas
mediante varios tipos de transporte grueso:

{kind=link}