 |
|
REACCIÓN ACROSÓMICA Y FECUNDACIÓNContenidos
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| TABLA 6 | |
|
Ovocitos fecundados (%) |
Ovocitos fecundados (%) |
|---|---|
|
88,5 ± 9,5 |
96,0 ± 4,0 |
Muestras de alrededor de 100 ovocitos con y sin espuma se inseminaron con 0,5 ml de una suspensión de espermatozoides (10 6 esp. / ml) |
| n=2 |
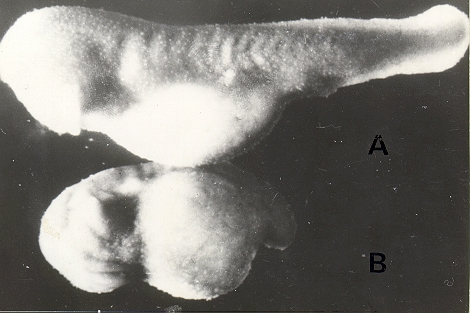 |
| Fig. 9: Embriones de Leptodactylus
chaquensis resultantes a partir de gametos provenientes de los
mismos ejemplares. Superior: criado en presencia de espuma. Inferior: criado en ausencia de espuma. |
La Tabla 6 muestra que la fecundación puede realizarse sin inconvenientes en ausencia de espuma. Sin embargo, los ovocitos fecundados en estas condiciones manifiestan anomalías en su desarrollo posterior
(Fig. 9). Algunos ensayos complementarios mostraron que la restitución de la espuma después de la fecundación previenen estas anomalías, permitiendo un desarrollo normal.
Los huevos de Leptodactylus chaquensis , a diferencia de otras especies, requieren oxígeno durante los primeros estadios de desarrollo. En efecto cuando se los incuba en condiciones de estricta anaerobiosis, el desarrollo queda bloqueado en los primeros estadios de la segmentación
(![]() ). Es probable, por lo tanto, que las anomalías observadas cuando se mantienen los ovocitos en ausencia de espuma sean debidas a una deficiente oxigenación. De ser correcta esta hipótesis una de las principales funciones de la espuma, en esta especie, sería la de asegurar la oxigenación del huevo en desarrollo. Por otra parte la falta de efecto de la ausencia de espuma en la fecundación, concuerda con la observación de que es posible fecundar a los ovocitos en condiciones de estricta
anaerobiosis (
). Es probable, por lo tanto, que las anomalías observadas cuando se mantienen los ovocitos en ausencia de espuma sean debidas a una deficiente oxigenación. De ser correcta esta hipótesis una de las principales funciones de la espuma, en esta especie, sería la de asegurar la oxigenación del huevo en desarrollo. Por otra parte la falta de efecto de la ausencia de espuma en la fecundación, concuerda con la observación de que es posible fecundar a los ovocitos en condiciones de estricta
anaerobiosis (![]() ).
).
Respecto al manejo de los gametos, cabe agregar que la viabilidad de los ovocitos en solución salina cae a medida que aumenta su tiempo de permanencia en la misma
(Tabla 7). Otro tanto se observa en el caso de los espermatozoides
(Tabla 8).
|
Tiempo (minutos) |
Ovocitos fecundados(%) |
|---|---|
|
0 |
100 ± 0,0 |
|
2,5 |
69,0 ± 31,0 |
|
5 |
62,5 ± 19,5 |
|
10 |
14,0 ± 3,0 |
|
15 |
8,0 ± 1,0 |
|
30 |
2,5 ± 2,5 |
|
|
|
n=2 |
Incubación de los espermatozoides de Leptodactylus chaquensis en solución salina y fecundación
|
Tiempo de incubación (minutos) |
Ovocitos fecundados |
|---|---|
|
0 |
75 |
|
15 |
5 |
|
30 |
0 |
|
45 |
0 |
|
|
Para este estudio nos hemos basado en el hecho ya mencionado de que el estado del acrosoma depende del medio en el que se hallan suspendidos los espermatozoides. La prueba consistió en inseminar ovocitos con espermatozoides provenientes de distintos medios, controlando la morfología del acrosoma, y la proporción de ovocitos fecundados.
Relación entre la morfología del acrosoma del espermatozoide de Leptodactylus chaquensis y fecundación
|
Solución |
Tiempo (min.) |
Fase 1 |
Fase 2 |
Fase 3 |
Fase 4 |
Fase 5 |
Motilidad |
Ovocitos |
|---|---|---|---|---|---|---|---|---|
|
Sacarosa 0,25M |
0 |
96 |
0 |
0 |
0 |
4 |
+ |
89,5% |
|
Sacarosa 0,25M |
5 |
98 |
0 |
0 |
0 |
2 |
+ |
90% |
|
Sacarosa 0,25M |
10 |
95 |
0 |
0 |
0 |
5 |
+ |
86,0% |
|
Sacarosa 0,25M |
30 |
92 |
0 |
0 |
0 |
8 |
+ |
87% |
|
Sacarosa 0,025M |
0 |
52 |
31 |
1 |
0 |
1 |
+++ |
61% |
|
Sacarosa 0,025M |
5 |
17 |
68 |
3 |
0 |
13 |
+++ |
2,5% |
|
Sacarosa 0,025M |
10 |
4 |
69 |
12 |
0 |
15 |
+ |
0 |
|
Sacarosa 0,025M |
30 |
4 |
53 |
25 |
0 |
18 |
- |
0 |
Ovocitos maduros en grupos de 200-300 se mantuvieron en cámara húmeda hasta su inseminación. |
| Las suspensiones de espermatozoides (106 esp. / ml) se prepararon en solución de sacarosa 0,25 M ó 0,025 M y se incubaron a 25 ± 2 °C. Alícuotas de 0,5 ml se utilizaron para inseminar los ovocitos luego de 0-5-10 y 30 minutos de su preparación . La motilidad y las fases del acrosoma se controlaron en alícuotas tomadas en el momento de la inseminación (otros detalles en Tabla 1 ). Cinco minutos después de la inseminación 2,5 ml de Holtfreter 0,1 fueron agregados a cada muestra. |
Los datos de la Tabla 9 muestran una correlación directa significativa entre el porcentaje de espermatozoides en fase 1 y el porcentaje de ovocitos fecundados (r: 0,998, p<
0,001). Este dato nos indica que para obtener altos porcentajes de fecundación es necesario partir de suspensiones de espermatozoides en las que no haya tenido lugar una reacción acrosómica espontánea. Quedaba por establecer, sin embargo, si los cambios en el acrosoma son necesarios para que el espermatozoide pueda penetrar las cubiertas de ganga que rodean al ovocito. Las observaciones directas y los
registros cinefotográficos efectuados
(![]() ) mostraron que los espermatozoides pueden cumplir su tránsito a través de las cubiertas de ganga manteniendo intacto su acrosoma
(Fig. 10 ).
) mostraron que los espermatozoides pueden cumplir su tránsito a través de las cubiertas de ganga manteniendo intacto su acrosoma
(Fig. 10 ).
 |
| Fig. 10: Penetración del espermatozoide a través de las cubiertas de ganga en Leptodactylus chaquensis (Contraste de fases, x 600) |
| AC: acrosoma; ES: espermatozoide; GA: ganga; OV: ovocito |
Los resultados anteriormente expuestos indican que la capacidad fecundante de los espermatozoides de
Leptodactylus chaquensis se mantiene en tanto no se produzca la reacción acrosómica espontánea. Según hemos visto, el hecho de que la
misma se produzca o no, parece depender de la tonicidad de las soluciones en que se suspenden los espermatozoides. La capacidad de ciertas soluciones para preservar la viabilidad de los espermatozoides ha sido reconocida también en otras especies de anfibios. En
Xenopus laevis (![]() ) este efecto fue denominado "capacitación artificial". Sin embargo creemos que este termino no resulta adecuado ya que capacitación tal como se emplea este termino para los mamíferos (
) este efecto fue denominado "capacitación artificial". Sin embargo creemos que este termino no resulta adecuado ya que capacitación tal como se emplea este termino para los mamíferos (![]() ), implica cambios en el espermatozoide que hacen posible la reacción acrosómica y la fecundación. Nuestros resultados más bien indican que se trataría de un efecto osmótico capaz de estabilizar las membranas del
acrosoma previniendo de esta manera la ocurrencia de una reacción prematura.
), implica cambios en el espermatozoide que hacen posible la reacción acrosómica y la fecundación. Nuestros resultados más bien indican que se trataría de un efecto osmótico capaz de estabilizar las membranas del
acrosoma previniendo de esta manera la ocurrencia de una reacción prematura.
El hecho de que no son necesarios cambios en el acrosoma del espermatozoide para atravesar las cubiertas de ganga, indican que el contenido del acrosoma seria utilizado a otro nivel, lo que será considerado en una sección aparte.
[Arriba]![]()
![]()
![]()
|















