 |
|
SECRECIONES DEL OVIDUCTO Y ACCIÓN DE LAS PROTEINASAS: SU RELACIÓN CON LA FECUNDACIÓN
Barra de Exploración
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 |
|
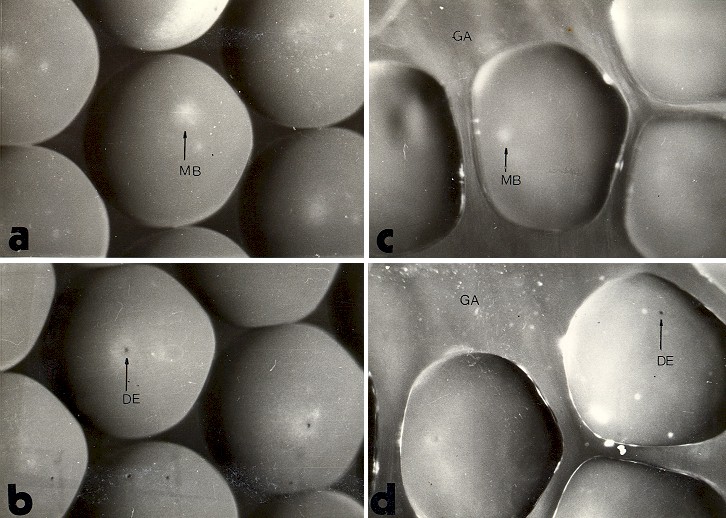
Fig 15: Ovocitos de cavidad y ovisaco con similares estadíos de maduración . |
Inseminaciones sucesivas de ovocitos uterinos mostraron que, considerando como punto inicial el momento de la inyección hipofisaria, el resultado obtenido variaba con el tiempo. Para tiempos muy cortos la mayoría de los ovocitos no presentan signos externos de activación, es decir que no rotan, ni la membrana vitelina se separa de la superficie del ovocito. A medida que transcurre el tiempo se observa que los ovocitos rotan pero la mayor parte de ellos no segmenta.
Más adelante los ovocitos rotan y segmentos aunque muchos de ellos no llegan al periodo de gastrulación. Finalmente, luego de transcurridas mas de 6 horas 30 minutos, el 100% de los ovocitos consigue superar esta etapa. La Tabla 25 muestra como van apareciendo estos fenómenos en una experiencia tipo. La
Fig. 16 muestra el aumento de los porcentajes de ovocitos fecundados a medida que transcurre el tiempo desde la inyección hipofisaria.
|
Fenómenos que acontecen en el ovocito en función del tiempo transcurrido desde la inyección hipofisaria |
|||
|---|---|---|---|
|
Fenómenos |
|||
|
|
Rotación |
Segmentación |
Gastrulación |
| 5 y 15 Hs. |
17 |
0 | 0 |
| 5 y 45 Hs. | 99 | 64 | 52 |
| 6 y 15 Hs. | 100 | 100 | 99 |
| 7 y 15 Hs. | 100 | 100 | 100 |
| 7 y 45 Hs. | 100 | 100 | 100 |
|
La experiencia se realizo con un ejemplar hembra de Bufo arenarum
conservado a 7 °C. Doce horas antes del experimento se llevo a 25 ± 2 °C. La
obtención de los ovocitos se realizo de la manera usual y se conservaron en
cámara húmeda a 25 ± 2 °C. |
|||
 |
| Fig. 16: Fecundidad de los ovocitos en función del tiempo de
tratamiento con hormonas hipofisarias. Se trabajó con ejemplares hembras de Bufo arenarum aclimatadas a 25 ± 2 °C. La obtención de los ovocitos se realizó de la manera usual. Los mismos se mantuvieron en cámara húmeda y grupos de alrededor de 100 ovocitos se inseminaron sucesivamente con espermatozoides provenientes de un mismo animal. La figura representa el porcentaje de ovocitos fecundados (M ± EEM, n=5) en función del tiempo transcurrido desde la inyección hipofisaria. |
La observación de la morfología externa de los ovocitos indica que entre las 4 horas 30 minutos y 5 horas, la mayor parte de ellos muestran en el polo animal una zona depigmentada en el centro
(Fig.
15: a y c) en la cual es posible observar a veces la mancha oscura que corresponde al huso de la primera división meiótica.
Entre las 5 y 5.30 horas, en la mayor parte de ellos se reduce el área depigmentada y en el centro de la misma aparece la fosa por donde saldrá el primer polocito. Mas allá de las 6 horas la mayoría ya expulsó el primer glóbulo polar
(Fig. 15, b y d).
Nuestros resultados establecen una relación estrecha entre el tiempo que transcurre desde la inyección hipofisaria y las etapas alcanzadas por los ovocitos uterinos. Este tiempo es necesario para que actúe sobre el protoplasma el material vertido desde la vesícula germinal y se produzcan, las modificaciones fisiológicas y estructurales que habilitarán al ovocito para su
desarrollo.
Los resultados obtenidos son coincidentes con los de otros autores en el sentido de que es necesario que transcurra un determinado tiempo para que el ovocito este en condiciones de ser fecundado
(![]() ), la única variación que existe es que este tiempo cambia en las diferentes especies de anfibios.
), la única variación que existe es que este tiempo cambia en las diferentes especies de anfibios.
Un punto importante de señalar, es que cuando se somete a la acción de las proteinasas acrosómicas ovocitos degangados y de cavidad, el efecto sobre la membrana vitelina de los primeros es mucho mayor que sobre la de los segundos. En efecto, las proteinasas acrosómicas actúan tardíamente sobre la membrana vitelina de los ovocitos de cavidad.
 |
 |
 |
| Fig. 17: Efecto lítico de los
espermatozoides sobre ovocitos de cavidad y de ovisaco 1: ovocito de cavidad; 2 ovocitos de ovisaco degangados. a: en ausencia de espermatozoides. b: 10 minutos después de agregar espermatozoides. c: aproximadamente 3 horas después de agregar los espermatozoides. DE: depresión que contiene el primer polocito. |
||
La Fig. 17.a muestra cuatro ovocitos: dos degangados y dos de cavidad, el aspecto externo es semejante. Sin embargo, luego de agregarse espermatozoides al medio puede observarse que las membranas de los ovocitos degangados sufren un claro efecto lítico, mientras que las de los de cavidad muestran escasas señales de ataque
(Fig. 17 : b y c). Este hecho confirma parte de la hipótesis enunciada, es decir la diferencia de susceptibilidad entre las membranas vitelinas de los ovocitos de cavidad y los uterinos. Sin embargo, el hecho de que para degangar ovocitos es necesario utilizar productos tales como el KCN o tioglicolato, cuyo efecto sobre la membrana ha sido señalado plantea la posibilidad de un artificio en estos resultados.
La primera porción del oviducto de Bufo arenarum, denominada pars recta no produce ganga, sino que en ella se acumula una secreción líquida. Esta porción representa la única parte del oviducto en la cual la membrana vitelina del ovocito se pone en contacto directo con las secreciones del mismo y, además, la única, a partir de la cual pueden obtenerse ovocitos sin ganga, sin necesidad de recurrir a ningún tratamiento químico. Resultaba por lo tanto, la porción ideal para comprobar la validez de los resultados anteriores.
Ensayando el efecto de las proteinasas acrosómicas sobre los ovocitos de esta porción de oviducto se observó que entre ellos y los de cavidad existían las mismas diferencias que entre los degangados y los de cavidad respecto a la sensibilidad a las proteinasas, es decir los ovocitos de pars recta son rápidamente atacados por las mismas.
Efecto del homogenado de pars recta
El pretratamiento de ovocitos de cavidad con homogenados de pars recta obtenidos a partir de animales estimulados con hipófisis condiciona a los mismos para que al ser tratados con proteinasas acrosómicas sus membranas sean atacadas rápidamente.
Fecundación de ovocitos de pars recta
Si se inseminan ovocitos tomados de la pars recta y de la cavidad del cuerpo en las condiciones señaladas en la Tabla 26 , los resultados indican que los ovocitos de la pars recta se fecundan en porcentajes significativamente superiores a los tomados de la cavidad del cuerpo.
| TABLA 26 | |
|
Inseminacion de ovocitos obtenidos de la cavidad celómica y de la pars recta del oviducto |
|
|
Origen de los ovocitos |
Ovocitos fecundados ( % ) M± EEM |
|
Pars recta |
84,8 ± 5,5 |
| Cavidad | 0,8 ± 0,8 |
|
Ovocitos maduros obtenidos de la pars recta y de la cavidad del cuerpo se colocaron en 2,5 ml de Holtfreter 0,1 y se inseminaron con 0,5 ml de una suspensión de espermatozoides ( 1 10 8 esp. /ml ) |
|
Fecundación de ovocitos de cavidad pretratados con pars recta
El pretratamiento de ovocitos de cavidad por medio de homogenado de pars recta preparado a partir de animales tratados con hipófisis, condiciona a los mismos para que al ser inseminados se obtengan altos porcentajes de fecundación (Tabla 27). Por el contrario cuando el pretratamiento se realiza con oviductos no estimulados este efecto no se observa.
| Tabla 27 | |
|
Efecto del homogenado de pars recta en la fecundación de los ovocitos de cavidad |
|
|
Pretratamiento |
Ovocitos fecundados ( % ) M± EEM |
| Holtfreter | 0,0 ± 0,0 |
| Homogenado de pars recta | 66,0 ± 15,1 |
|
Ovocitos maduros obtenidos de la cavidad del cuerpo fueron incubados en Holtfreter standart y homogenado de pars recta durante treinta minutos. Las muestras se lavaron tres veces con 50 ml de Holtfreter 0,1 y se inseminaron. El medio de inseminación contenía 2,5 ml de Holtfreter 0,1 y al mismo se agregaron 0,5 ml de una suspensión de espermatozoides (1.108
esp / ml). |
|
Durante las experiencias se realizaron análisis cariológicos de gástrulas (obtenidas al azar). Las mismas mostraron cariotipo normal. Por otra parte tomando las debidas precauciones, el desarrollo se prolonga hasta la metamorfosis.
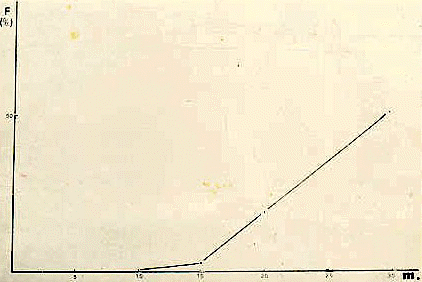
La única diferencia apreciable entre la inseminación de los ovocitos condicionados con pars recta y los ovocitos con ganga, es un retardo en la aparición del primer surco. Esta diferencia puede ser explicada en base al retardo en la velocidad de fecundación
(Fig. 18) ya que algunos datos obtenidos indican que los valores son bastante diferentes de los con ganga (
![]() ).
).
 |
|
Fig. 18: Velocidad de fecundación en ovocitos condicionados. |
Algunas propiedades del factor pars recta
El factor responsable del efecto biológico sobre los ovocitos es termolábil (Tabla 28) y no dializable.
| Tabla 28 | |
|
Efecto de la temperatura sobre el homogenado de pars recta |
|
|
Pretratamiento |
Ovocitos fecundados ( % ) |
|
Homogenado de pars recta |
53 |
|
Homogenado de pars recta inactivado por calor |
0 |
|
Una alícuota de homogenado de pars recta se inactivo durante 5 minutos en agua hirviendo. La prueba de actividad se realizo sobre ovocitos de cavidad en las condiciones de la Tabla 27. |
|
Localización del factor pars recta
Si con las técnicas inmunológicas corrientes se preparan anticuerpos contra homogenados de pars recta , los anticuerpos generados inhiben el efecto de dicho homogenado sobre los ovocitos. En efecto, tal como se puede observar en la
Tabla 30, si ovocitos previamente condicionados con pars recta son tratados con suero anti pars recta se observa una significativa inhibición de los porcentajes de fecundación. Los controles realizados con suero de animales no tratados no presentaron inhibición. Por otra parte, el tratamiento de los ovocitos cavitarios con homogenados de pars recta conjugados con isotiocianato de fluoresceína permitió localizar la fluorescencia en la membrana vitelina (![]() ).
).
|
Accion del suero anti pars recta sobre la fecundidad de los ovocitos condicionados |
|
|
Pre - incubación |
Ovocitos fecundados ( % ) M ± EEM |
| Suero normal | 46,4 ± 2,6 |
| Suero anti pars recta | 0,0 ± 0,0 |
|
Ovocitos condicionados se trataron con suero normal y suero anti pars recta durante 2 horas. Luego del tratamiento, los ovocitos se lavaron con Holtfreter 0,1 y se inseminaron en condiciones similares a las descriptas en la Tabla 25. |
|
El conjunto de las observaciones descritas en esta sección nos induce a suponer que la modificación de la membrana vitelina o su "maduración" por los factores del oviducto (para emplear el termino acuñado por Schuetz) es uno de los pasos que sigue el ovocito para asegurar se fecundación. Poco conocemos acerca del mecanismo de acción de este factor, las evidencias con anticuerpos y homogenado marcado indican que el mismo se pega a la membrana vitelina pero desconocemos todavía si un mecanismo enzimático está envuelto en este proceso.
![]()
![]()
![]()
|