|
 |
|
TEMA 23
|
|
|
Tema 23:
Reproducción y Polinización
|
MEGASPOROGÉNESIS y MEGAGAMETOGÉNESIS
Gimnospermas
Ver
en Animaciones el Ciclo vital del Pino
|
Ciclo vital de Pinus |
 |
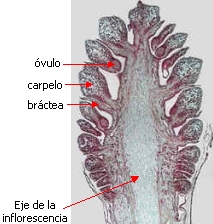
Las inflorescencias
femeninas de Pinus, los conos o piñas, son de mayor tamaño y mucho más complejas que las
inflorescencias masculinas.
Las brácteas se disponen helicoidalmente alrededor del eje de la
inflorescencia o estróbilo. Cada flor
femenina está inserta en la axila de una bráctea
tectriz. La flor es desnuda, está
constituída solamente por un carpelo o escama ovulífera.
Cada escama ovulífera
(carpelo) lleva 2 óvulos en su cara superior;
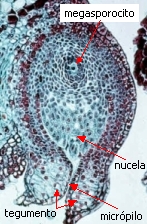
cada óvulo consiste de la nucela rodeada por un
tegumento y con el micrópilo orientado hacia el eje de la
inflorescencia. La nucela es el equivalente del megasporangio.
Megasporogénesis en Pinus
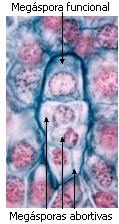
En cada megasporangio se diferencia una sola célula madre de
las megásporas o megasporocito. Después de la polinización el
megasporocito se divide por meiosis, formando una tétrade
lineal de megásporas. Las tres orientadas hacia el
micrópilo abortan, y sólo la ubicada más profundamente es
funcional
Megagametogénesis en Pinus
La megáspora funcional se divide por mitosis muchas veces,
iniciando la formación del gametofito femenino. Inicialmente hay una etapa de divisiones nucleares libres, sin formar paredes celulares, hasta constituir un
cenocito con
unos 2000 núcleos. En este estado pasa el invierno, y en la
primavera siguiente reanuda el crecimiento.
Entonces comienza la
formación de paredes entre los núcleos del cenocito,
constituyendo el endosperma primario o prótalo o gametófito
femenino. |
Pinus: óvulos con gametófitos femeninos
en formación |
|
En divisiones nucleares
libres |
Inicio de la formación
de paredes celulares |
|

|
 |
|
http://departments.bloomu.edu/biology/pics/botany/B500w.jpg |
Una vez que termina la formación de paredes
celulares, en el prótalo se forman 2 o 3 arquegonios hacia el extremo
micropilar. Cada arquegonio está formado por una ovocélula
o gameto femenino voluminoso,
por encima del cual están las células del canal del cuello y una célula
del canal del vientre ubicada en el centro. El
gametófito femenino maduro es el prótalo con los arquegonios.
|
Óvulos de |
Gametofito femenino
maduro |
Detalle de los arquegonios |
 |
 |
|
http://www.uleth.ca/bio/ bio1020/images/ |
http://departments.bloomu.edu/biology/pics/botany/ |
Angiospermas
Ver
en Animaciones el Ciclo vital del Paraíso
|
Ciclo vital de Melia
azedarach, Paraíso, angiosperma. |
 |
En las angiospermas, la parte femenina del proceso reproductivo sexual tiene lugar en los
óvulos. Cada óvulo está inervado por un
haz vascular
que atraviesa el funículo y llega hasta la cálaza;
en los óvulos anátropos recorre toda la rafe.
En raras oportunidades el haz vascular se ramifica e inerva
parcial o totalmente los tegumentos, como ocurre
en los óvulos de algunas Annonaceae, Euphorbiaceae y en los del irupé.
|
Vascularización de
óvulos anátropos |
|
Linum, haz
vascular en la rafe |
Artabotrys
(Annonaceae):
tegumento vascularizado |
Victoria cruziana,
irupé, esquema tridimensional |
 |
 |
 |
|
Esquema: Svoma 1997 |
Esquema: Valla &
Martin 1976 |
La nucela
del óvulo es el megasporangio.
En la nucela se diferencia una célula
madre de las megásporas o megasporocito.
En los óvulos
tenuinucelados la célula está en posición subsuperficial,
debajo de la epidermis. En los óvulos
crasinucelados, se diferencia profundamente en la
nucela.
|
Spigelia
humboldtiana: óvulo
tenuinucelado |
Lopezia: óvulo crasinucelado |
|

|
 |
|
Imagen de Maldonado 1989 |
Imagen
de ??? |
Megasporogénesis en Angiospermas
|
Síntesis de la megasporogénesis y
megagametogénesis en Angiospermas |
 |
El megasporocito se divide por meiosis formando
cuatro megásporas
haploides que se disponen en una tétrade
lineal. Comúnmente las tres ubicadas hacia el micrópilo, degeneran, y la
más interna originará el saco embrionario o
gametófito femenino. A
menudo se forma una pared de calosa
durante la meiosis, que aisla la megáspora que sobrevive (en
los sacos tetraspóricos no se forma calosa).
|
Megasporogénesis en Lilium,
angiosperma, en transcortes de ovario |
|
Lóculo con 2 óvulos |
Detalle del ápice de
un óvulo |
Óvulo con tétrade
lineal de cuatro megásporas |
 |
 |
 |
|
Imágenes de http://images.iasprr.org/lily/female.shtml |
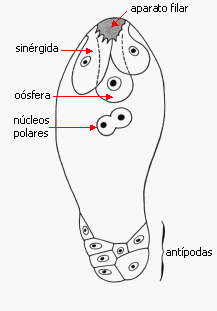
Megagametogénesis en Angiospermas
|
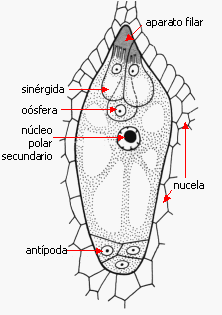
Las sinérgidas son las células más llamativas por su
organización: son células de transferencia, presentan en el
ápice el aparato filar, una pared con una serie de
protuberancias internas, de apariencia fibrosa, formada por
hidratos de carbono, hemicelulosa y pectina principalmente.
Su
función es la de atraer y recibir al tubo polínico, y también
está involucrada a menudo en absorber nutrientes de la nucela y
en hacerlos llegar a la ovocélula. En algunos casos forman
haustorios.
La pared que las separa de la oósfera a menudo es
incompleta, hacia el polo calazal a menudo hay solo plasmalema.
|
Arabidopsis thaliana:
sinérgidas
sc:
sinérgidas; fa:
aparato filar
cw: pared celular;
|
|

|
|
Imagen de Kasahara et al.
2005 |
El grupo que se ubica hacia el polo calazal del saco
embrionario constituye las antípodas
que aparentemente participan en la nutrición del saco
embrionario; en ciertas especies son haustoriales. En las gramíneas
habitualmente se encuentran varias antípodas, en Sasa
paniculata, bambú, hay unas 300 antípodas en cada saco. Los
dos núcleos restantes, denominados núcleos polares se
ubican en la célula del medio, y frecuentemente se fusionan antes
de la penetración del tubo polínico, constituyendo el
núcleo secundario 2n Tipos de saco embrionario
Hay varios tipos de sacos
embrionarios según el número de macrósporas que intervienen en
su formación, y el número de divisiones mitóticas que se
producen.
El tipo más común (80% de los casos conocidos) es el
MONOSPORICO, que deriva de una macróspora. Es el tipo
fundamental, llamado tipo Polygonum porque se descubrió en este
género.
Una variante es el saco monospórico tipo Oenothera: la macróspora sufre
sólo dos divisiones dando 4
células, 3 forman el aparato ovular (la oósfera y 2
sinérgidas) y la cuarta es la célula del
medio, que en este caso es haploide. |
Melia azedarach, paraíso:
saco embrionario tipo Polygonum
|
 |
SACOS BISPORICOS. Intervienen 2
macrósporas en su formación. En el tipo Allium, cada macróspora
sufre 2 mitosis, y los 8 núcleos resultantes se ubican igual que
en Polygonum.
SACOS TETRASPORICOS. Participan las
4 megásporas en la formación del saco embrionario. En el tipo Adoxa: cada megáspora se divide una vez, y
los 8 núcleos se acomodan como en Polygonum.
En el saco tetraspórico
tipo Fritillaria: 3 megásporas se fusionan formando un
núcleo 3n; las células sufren dos divisiones, se forman 8
núcleos, cuatro n, y cuatro 3n. Las células del aparato ovular y un nucleo
polar son n; las antípodas y un núcleo polar son 3n; el
núcleo secundario de la célula del
medio es 4n. El género Lilium presenta
este tipo de saco embrionario.
Es importante señalar que no es raro hallar
más de un tipo de saco embrionario en una especie o incluso en
el mismo individuo. En Delosperma (Aizoaceae) se hallaron 5
tipos diferentes (Mauseth, 1988). En
el saco embrionario de Plumbago no hay sinérgidas, la ovocélula
presenta aparato filar en el polo micropilar.
|
Tipos de Saco
Embrionario |
 |
POLINIZACIÓN
 |
La
polinización es el transporte de los granos de polen
desde los sacos polínicos de las anteras hasta el
micrópilo de los óvulos en Gimnospermas y
hasta el estigma en las Angiospermas.
La Biología floral o
ecología floral es la ciencia que se ocupa de investigar la
variedad de fenómenos que se producen en la polinización de
los espermatófitos.
|
Cleistogamia y casmogamia
La polinización puede producirse antes o después
de la antesis. El primer caso es la cleistogamia,
cuando la polinización se realiza ya en el capullo o botón
floral, la autogamia o sea la fecundación con las gametas del
propio polen, es obligada
porque las flores no se abren.
Su ventaja es que permite, a una especie
bien adaptada, perpetuarse en un medio más o menos estable.
Su desventaja es que por la autogamia, la especie presenta menor variabilidad hereditaria,
y pierde plasticidad evolutiva.
Hay plantas como el maní, Arachis (Leguminosa), y la violeta, Viola
odorata (Violaceae), que presentan
los dos tipos de flores: producen flores
casmógamas, y al
comienzo o al final de la floración presentan flores
cleistógamas, de tamaño, forma y color diferentes. En Viola
odorata las flores cleistógamas
tienen pétalos rudimentarios, y las
anteras son más pequeñas, con menor cantidad de polen.
La casmogamia es el proceso que ocurre
después de la antesis, en flores abiertas. En las flores
casmógamas puede tener lugar la autogamia o la alogamia,
polinización cruzada.
POLINIZACION DIRECTA - AUTOGAMIA
Cuando el transporte de polen, y por ende, la fecundación,
ocurre entre flores del mismo individuo, el proceso
se denomina autogamia. Está muy difundida entre
las malezas, las plantas pioneras y las especies insulares, que
necesitan la fructificación de individuos aislados. En especies
autógamas, las flores con frecuencia son inconspicuas, con piezas florales
reducidas, menor cantidad de polen, sin fragancia y sin néctar.
| En las flores monoclinas o perfectas
o cosexuales, es posible la autofecundación, ya sea
por la acción de diversos dispositivos florales o por la
intervención de un polinizador.
En el lirio, Lilium martagon, el estilo
inicialmente erecto, se mueve curvándose para ponerse en contacto con los
estambres para autopolinizarse.
|
Lilium martagon:
estilo móvil |
 |
|
Foto de Spencer C.H.Barrett |
En la achicoria, Cichorium intybus, las flores son protándricas, es decir que
el androceo madura primero; el estilo al crecer,
se carga de polen en su cara externa. Si no ocurre polinización
cruzada por medio de insectos, las ramas estigmáticas se
alargan y se curvan sobre sí mismas, poniendo en contacto su
superficie receptiva interna con el propio polen.
Autopolinización en
Cichorium intybus, achicoria |
 |
|
Esquemas modificados de Valla
1979 |
| En Turnera orientalis, con
flores homostilas, cuando no hay polinización
cruzada por medio de insectos, ocurre la autogamia al marchitarse
la corola, porque los pétalos se juntan y retuercen poniendo en
contacto anteras y estigmas. Se forman frutos en ausencia
de polinizadores. |
Turnera orientalis,
flor homostila |
Flor marchita |
 |
 |
En flores erectas, disciformes y cóncavas, como
las de Ranunculus, el agua de lluvia puede provocar la
autopolinización o, por salpicadura, la alogamia.
Cuando la polinización ocurre entre flores
distintas del mismo individuo se habla de geitonogamia,
situación común en especies con floración masiva, como el
lapacho (Tabebuia heptaphylla), el ibirá-pitá (Peltophorum
dubium), el palo borracho (Ceiba speciosa). Genéticamente es equivalente a la
autogamia.
Si la planta es monoica, como la de maíz,
por ejemplo, la geitonogamia es
obligada.
| Peltophorum dubium,
ibirá-pitá |
Tabebuia heptaphylla,
lapacho |
Delonix regia,
chivato |
 |
 |
 |
POLINIZACIÓN CRUZADA - ALOGAMIA
Cuando el transporte de polen ocurre entre flores de individuos
diferentes, tenemos polinización cruzada, y por ende,
fecundación cruzada o alogamia. En
muchas especies es obligada, a veces las flores, aún cuando sean
cosexuales (hemafroditas), son autoincompatibles,
es decir que tienen barreras genéticas y fisiológicas que
impiden la germinación del propio polen o el desarrollo del
tubo polínico (Malus sylvestris,
manzano). La autoincompatibilidad puede ser esporofítica o
gametofítica:
 |
La incompatibilidad
esporofítica depende de la pared del grano de polen, que es de
origen esporofítico. Para que el grano de polen pueda
germinar, debe adherirse al estigma, lo que ocurre solamente
cuando hay compatibilidad entre las proteínas de reconocimiento
que se encuentran en la esporodermis, y los receptores que existen
en el estigma. |
|
La incompatibilidad gametofítica
depende
de la
constitución
genética del
gametofito
masculino, el polen puede germinar, pero
el crecimiento del tubo polínico es detenido después de
su penetración en el estilo. Esta in |
|
Autoincompatibilidad
esporofítica |
Autoincompatibilidad
gametofítica |
 |

|
|
Imágenes de http://www2.unil.ch/lpc/images/docu04/illustr_flor.htm |
Las ventajas de la alogamia radican en la producción de nuevas combinaciones
genéticas en la población, que aseguran la variabilidad de la especie
y en consecuencia, la
posibilidad de sobrevivir a los cambios de medio ambiente. Por
eso las Angiospermas desarrollaron numerosas adaptaciones
florales para favorecer la alogamia, como por ejemplo la
separación espacial y temporal de los sexos y otras variaciones
como la presentación secundaria de polen.
Las flores de las plantas que poseen un sólo tipo
de flores, frecuentemente presentan estambres y pistilo de
longitud semejante, y se denominan homostilas.
DICOGAMIA (separación temporal).
Los estambres y estigmas de una misma flor no alcanzan al mismo
tiempo la madurez
para la polinización. Si los estambres
maduran antes, las flores son protándricas, la flor funciona
primero como flor masculina y luego como flor
femenina. La protandria favorece la alogamia , y es el caso más
frecuente en especies con dicogamia intrafloral.
|
Protandria en Agave
(Bromeliaceae) |
|
Inflorescencia |
Inflorescencia parcial |
Flores
protándricas |
Flores en
estado femenino |
 |
 |
 |

|
| www.infojardin.com/ |
|
|
|
Cuando
el gineceo
madura antes que el androceo, las flores son proteroginas o protoginas (Magnolia
grandiflora, Plantago, especies de Leguminosas).
En este caso, la flor funciona primero como flor femenina
y luego como flor masculina. Es menos frecuente.
Frecuentemente la protoginia
intrafloral está asociada con autocompatibilidad, así que este
proceso parecería un recurso evolutivo para asegurar la producción de
frutos y semillas (Bertin & Newman 1993). |
Protoginia en Magnolia
grandiflora |
 |
 |
|
Imágenes de K.R.Robertson,
http://www.life.uiuc.edu |
|
La dicogamia puede ocurrir tanto en plantas con
flores monoclinas (intrafloral) como en plantas monoicas con
flores diclinas (interfloral). Por ejemplo, en Cucurbita
maxima var. zapallito, el zapallito tronco, con flores
diclinas, primero aparecen las flores masculinas y unas dos
semanas después, las flores femeninas. |
Cucurbita maxima,
flor femenina |
C. maxima, flor
masculina |
|

|
 |
HERCOGAMIA (separación espacial)
La
presentan aquellas flores en las cuales las anteras y estigmas están muy
separados unos de otros; es bastante común, generalmente reduce
la autopolinización intrafloral, tal como la dicogamia.
En algunos casos muy especializados las flores poseen una conformación tal que
dificulta o impide la autogamia (Iris, Asclepiadaceae,
Orchidaceae). Las flores de Ayenia (Sterculiaceae)
presentan androginóforo, que eleva el gineceo y el tubo estaminal
formado por 5 estambres y 5 estminodios. Los pétalos presentan
una larga uña, y cada uno se apoya sobre el tubo estaminal por
encima de un estambre. De esa manera, el estigma queda por
encima de los pétalos, y los estambres por debajo, impidiendo la
autopolinización
|
A. Flor de Ayenia
entera en vista súpero-lateral
B, vista lateral del androginóforo, tubo estaminal y un
pétalo |
Ayenia
mansfeldiana,
flor en vista lateral |
 |
 |
| Dibujos
de Cristóbal,1960.
Foto de
M.Bonifacino, http://www.Plantsystematics.org
Naked image |
Según la posición relativa de anteras y estigmas se
presentan varios tipos:
1) Hercogamia de aproximación:
los estigmas están por encima de las anteras, y son los
primeros en contactar a los polinizadores a su ingreso en la
flor.
2) Hercogamia revertida: las
anteras están por encima de los estigmas; ocurre en flores
tubulosas, polinizadas por lepidópteros.
|
Hercogamia de
aproximación en Turnera |
Hercogamia revertida |
 |

|
3) Hercogamia recíproca o
Heterostilia. Las poblaciones están compuestas por
individuos hermafroditas, pero hay 2 (distilia) o 3 (tristilia)
tipos de individuos con formas florales caracterizadas por la
disposición y longitud recíproca de anteras y estigmas.
En las especies distilas hay flores
longistilas (estilos largos y estambres cortos) y brevistilas
(las anteras están por encima de los estigmas). Los granos de polen
de las flores brevistilas son mayores, y a veces presentan
escultura diferente. En las flores con estigmas papilosos hay correspondencia entre el tamaño de los granos de polen y
la longitud de las papilas del estigma.
En las especies
tristilas hay una tercer forma floral, las flores mediostilas.
El estigma y el estilo actúan como filtros
impidiendo la germinación o llegada del polen propio:
autoincompatibilidad genética, hay barreras a distintos
niveles. La fructificación resulta óptima sólo cuando la
polinización es cruzada.
Se presenta con frecuencia en flores tubulosas,
actinomorfas, nectaríferas, polinizadas por diferentes
insectos. La distilia es frecuente en Sterculiaceae, Plumbaginaceae, Turneraceae,
Rubiaceae y Borraginaceae. La tristilia se presenta en los
géneros Lythrum, Pontederia, Eichhornia y Oxalis.
|
Distilia en Primula |
Distilia |
 |
|

|
|
Tristilia |
 |
Nasrallah y col. (1999) hallaron un gene localizado en el locus
S: "cysteine-rich protein," llamado SCR para abreviar.
Es el determinante masculino del proceso reproductivo de la
planta. El gene es la clave de porqué el estigma aceptará el
polen de otro pie o rechazará el polen del propio pie. El gene
se expresa en la antera, donde se produce y codifica el polen.
4) Hercogamia interfloral o
monoecia. Las plantas
monoicas producen flores unisexuales,
femeninas y masculinas. Este fenómeno evita la autopolinización; puede estar
asociada o no con autoincompatibilidad. Las plantas monoicas
autocompatibles presentan geitonogamia obligada.
Se ha
comprobado que las especies monoicas pueden alterar la proporción de
flores masculinas y femeninas en respuesta a estímulos
ambientales. Esta plasticidad reproductiva es la principal
virtud del sistema.
5) Enantiostilia. Es un
fenómeno por el cual el pistilo, órgano sexual femenino, está
desviado a la izquierda o a
la derecha, de manera que cada tipo de flor es la imagen
especular (mirror-image) de la otra. Esta asimetría promueve la
polinización cruzada en plantas visitadas por abejas, y
disminuye el nivel de geitonogamia. Se
presenta en flores sin néctar, con anteras dimorfas: la flor
presenta varias anteras que proveen polen a los insectos, y una
antera polinizadora. Cuando ambos tipos de flor
se encuentran en el mismo individuo, se denomina enantiostilia
monomórfica, presente por ejemplo en Cassia chamaecrista
y Solanum rostratum. Cuando cada tipo de flor está en individuos
diferentes se denomina enantiostilia dimórfica, por ej:
Cyanella alba (Jesson & Barrett 2003).
| Enantiostilia
en Cassia chamaecrista |
Tipos de
enantiostilia |
 |

|
| Foto de Jesson; esquemas de
Jesson & Barrett 2003 |
D: flor derecha. I:
flor izquerda |
6) Dimorfismo estigmático altitudinal
(Stigma-height dimorphism). Las plantas presentan dos
tipos florales que difieren en la altura de los estigmas. En la
forma L (longistila), los estigmas están ubicados por encima de
los estambres, mientras en la forma B (brevistila) los estigmas
están por debajo de las anteras. La diferencia con la
heterostilia es que la longitud de los estambres es similar en
los dos casos, o sea que no hay reciprocidad entre la longitud
de las estambres y de los pistilos de los dos tipos florales. Se
presenta en familias con heterostilia y sin heterostilia (Ericaceae, Liliaceae,
Amaryllidaceae) (Barrett & Harder 2005).
| Dimorfismo
estigmático altitudinal en Narcissus assoanus |
 |
 |
|
Esquemas y foto de Baker et
al. 2000 |
DIOECIA
Es la separación de los sexos en distintos pies (mamón,
palmera datilera, el género Baccharis (Compuesta) al que
pertenece la carqueja, B. articulata). Cada especie
presenta individuos con flores masculinas e
individuos con flores femeninas, lo que determina la alogamia obligada.
Muchas plantas dioicas presentan flores relativamente pequeñas,
blancas, amarillentas o verdosas, de morfología no
especializada, que atraen una variedad de insectos pequeños,
especialmente abejas pequeñas de las familias Halictidae,
Megachilidae y Meliponini. A menudo está asociada con plantas
de gran tamaño y polinización abiótica (Salix, sauce). La dioecia es rara en
plantas con flores grandes, especializadas, con morfología
compleja.
Baccharis
pingraea, planta femenina
|
 |
 |
|
Dibujos de Flora de
Entre Ríos |
PRESENTACIÓN SECUNDARIA DE POLEN.
Es la
transferencia del polen de las anteras a otra estructura de
donde es recogida por los polinizadores. Es virtualmente
universal en las Compositae y es común en las Leguminosae
Papilionoideae. Por ejemplo en Pisum sativum (arveja) el estilo
lleva sobre la cara adaxial un conjunto de pelos que semejan un
cepillo. La quilla y el estilo son reflejos. Con la visita de un
polinizador el estilo baja, y al retomar su posición recoge
polen sobre el cepillo desde el cual lo recoge el polinizador.
Pisum sativum,
arveja, flor con presentación secundaria del polen |
|

|
A. Vista frontal de la flor
B. Corte longitudinal de la flor
C. Pétalos separados:
1. estandarte o vexilo
2. alas
3. quilla
D. Androceo diadelfo y Gineceo
E. Gineceo, con estilo reflejo
y cepillo de pelos en su parte
superior |
 |
| Foto
de ??? |
En Stylosanthes gracilis los estambres son monadelfos; antes de la antesis las anteras pequeñas y
redondeadas de los estambres del verticilo interno están por
debajo de las anteras grandes, ya dehiscentes, de los estambres
del verticilo externo. Enseguida de la antesis las anteras
pequeñas están por encima de las grandes, sus filamentos se
han alargado rápidamente, y junto con el estilo ayudan a
empujar el polen hacia afuera cuando una abeja visita la flor.
En la antesis tardía, con la visita de otra abeja, se abren y
presentan el polen las anteras pequeñas (Yeo, 1993)
AGENTES POLINIZADORES
Los granos de polen son inertes, su transporte
está asegurado por agentes externos, abióticos como agua y
viento, o bióticos como animales diversos.
HIDROFILIA: Polinización por medio del agua.
Vallisneria (monocotiledónea acuática.) presenta flores
flotantes, las femeninas permanecen fijas a la planta por un
largo pedúnculo floral; las flores masculinas se desprenden,
flotan, y son llevadas por la
corriente del agua o el viento hasta las flores femeninas.
|
Vallisneria spiralis
(monocotiledónea) - polinización hidrófila |
 |
 |
| Foto
de ??? |
|
Algunas Angiospermas hidrófitas como
Potamogeton striatus y P.pusillus, Najas marina
(Monocotiledóneas)
Ceratophyllum demersum
(Dicotiledónea) propias de nuestros humedales, tienen flores
masculinas y femeninas sumergidas, en ese caso la reproducción también está adaptada
y la polinización es hidrófila. En todos los casos el polen
es llevado por el agua hasta los estigmas, y a pesar de que las
plantas no están emparentadas, el polen tiene aspecto similar:
es filamentoso, flexible, pegajoso. Las flores de Zostera
marina, planta que vive sumergida en el mar, las flores presentan granos de
polen filiformes de más de 2 mm de longitud, y las unidades se
adhieren entre sí formando copos. En otras especies con
polen esférico, las
unidades van embebidas en largas tiras de mucílago. Su forma
facilita el contacto y la adherencia a los largos estigmas.
| Zostera marina,
planta sumergida y polen y estigma |
 |
 |
| http://www.laisladelsur.com |
Imagen de Cox 1993 |
ANEMOFILIA: Polinización por medio del
viento
La polinización por medio del viento se presenta en la mayoría de las Gimnospermas.
Es más frecuente en Monocotiledóneas (Gramíneas, Cyperáceas,
Palmeras)
que en Dicotiledóneas (Salicáceas, Chenopodiáceas, Fagáceas). El
transporte de polen no está orientado, por lo cual se producen
grandes cantidades de polen, de tamaño pequeño, superficie
lisa (facilita la dispersión), y seco, por escasa formación de
cemento polínico o por su rápida desecación.
En Pinus (Gimnosperma) los granos de polen
tienen sacos aeríferos para aumentar la flotabilidad, llegan
directamente a los óvulos, que presentan en el micrópilo gotas
receptoras de polen, mucilaginosas o azucaradas. En ese momento,
las escamas y las brácteas de la inflorescencia están bien
separadas. Cuando la gota de fluído micropilar se evapora, el
polen se hunde y queda sobre la nucela. Después de la
polinización las escamas se juntan, protegiendo a los óvulos
entre ellas. Poco después que el polen queda en contacto con la
nucela, germina, emitiendo el tubo polínico.
En Angiospermas las flores anemófilas carecen
de medios de atracción (perianto, olor, néctar), suelen ser
unisexuales, las masculinas más numerosas que las femeninas,
que generalmente son uniovuladas. Los estilos y estigmas están
agrandados para facilitar la captación del polen, como se puede
ver en las flores de Poterium
sanguisorba; la expulsión del polen está facilitada por la
movilidad de los filamentos estaminales (Gramíneas) o del eje de la inflorescencia (gimnospermas,
Quercus, Alnus). La floración es temprana, las flores aparecen
antes que el follaje que puede obstaculizar la captación del
polen. Las plantas están reunidas en poblaciones grandes y en
lugares expuestos al viento (llanuras o estratos superiores de
los bosques). No hay plantas anemófilas en las selvas
tropicales ricas en animales antófilos.
La anemofilia tiene baja eficiencia. Se
calcula que un pie de maíz produce 50 millones de granos de
polen; para fecundar los óvulos de un pie son necesarios sólo
1000 granos.
Paspalum dilatatum,pasto miel (Gramínea),
inflorescencia muy
aumentada |
Alnus acuminata,
aliso,
amentos masculinos péndulos |
Poterium
sanguisorba:
flores con polinización anemófila (muy aumentadas) |
 |
 |
 |
|
http://www.nybg.org |
Dibujos de Strasburger |
ZOOFILIA: Polinización por medio de
animales.
Agentes polinizadores variados, los más
comunes insectos (Entomofilia), pájaros (Ornitofilia) o
murciélagos (Quiropterofilia). Los agentes polinizadores buscan
alimentos, recompensas, que son el polen (rico en proteínas,
grasas, glúcidos y vitaminas) o el néctar. Las flores
presentan atractivos para asegurar la visita de los agentes, que
pueden ser de naturaleza óptica (color) o química (olor).
Los olores atraen a los polinizadores, algunos
tienen el olfato muy desarrollado, especialmente los insectos y
los murciélagos. De acuerdo con la sensibilidad humana, los
olores se clasifican en simpáticos (agradables) o idiopáticos
(desagradables). Entre los primeros tenemos olores
aromáticos (canela, vainilla, etc.), dulces (miel, rosa, violeta,
etc.), a frutas (naranja, banana, etc.). Entre los olores
idiopáticos tenemos olores fétidos (olor a carne en
descomposición, a excrementos) y olores feos (a pescado, caprino,
etc.). También están las feromonas, aromas producidos por
los animales para provocar una respuesta de comportamiento
reproductivo en un receptor de la misma especie.
Raramente todas las especies de una familia presentan el mismo
tipo de polinización: las Aristolochiaceae son uniformemente miófilas y las Loranthaceae, Cannaceae y Heliconiaceae son casi
exclusivamente ornitófilas. En general en cada familia o en el
mismo género hay especies adaptadas a diferentes polinizadores
(Vogel 1990).
Flores Entomófilas,
polinizadas por insectos.
La diversificación de las
Angiospermas y de los
insectos se produjo al mismo tiempo (co-evolución).
El polen de
las flores entomófilas presenta púas, espinas, irregularidades
que facilitan la adhesión, cemento polínico o hebras de viscina.
Si los polinizadores transportan el polen en el
dorso, se habla de nototribia, si lo llevan sobre la cara
ventral, de esternotribia. |
Polen de plantas
entomófilas (MEB)
|
 |
Coleópteros: flores
cantarófilas. Son las más primitivas, son flores poliníferas,
es decir que ofrecen polen como recompensa, tienen
muchos estambres que producen polen en exceso. Son generalmente
flores rotáceas, fácilmente accesibles, robustas, verdosas o
blanquecinas, fuertemente olorosas. Son frecuentes en las
Magnoliaceae, Ranunculaceae, Nymphaeaceae. Probablemente las Rosaceae,
Papaveraceae y Dilleniaceae (Paeonia) son secundariamente poliándricas y poliníferas
.
Los coleópteros, con sus fuertes mandíbulas
masticadoras, además de comer las anteras, destruyen frecuentemente diversas partes
florales, por esta razón los óvulos de las flores cantarófilas
están profundamente ubicados.
| Flores
cantarófilas: Nuphar (Nymphaeaceae) y Hepatica
americana (Ranunculaceae) |
|

|
 |
| Imagen
de ???? |
Foto de Raven 2003 |
La inflorescencia de muchas Aráceas, cuando está
lista para la polinización, sufre un brusco aumento de
temperatura, gracias a la oxidación de sustancias de reserva
acumuladas (en Philodendron scandens, planta nativa de
América del Norte, la inflorescencia alcanza 46° cuando la
temperatura ambiente es de 4°). Los coleópteros que visitan
estas plantas son atraídos por el olor desagradable, similar a
la carne en putrefacción, provocado por sustancias químicas
como aminas e indoles, que se dispersan más eficientemente con
el calor.
Al atardecer, se reunen cantidades de coleópteros en
el interior de la espata, y allí se alimentan de polen y partes
florales al tiempo que se aparean.
Al otro día salen cargados
de polen, que depositan sobre las flores femeninas de la
inflorescencia en la que se alojan la noche siguiente |
Philodendron (Araceae) |
 |
Dípteros (moscas): flores Miófilas.
Las flores polinizadas por estos insectos son heterogéneas.
| Algunas flores son más o menos inodoras,
presentan corola pequeña y
néctar libre. Una especie polinizada por moscas es Cabomba
caroliniana, planta acuática de nuestros
humedales.
Cada flor de 2 cm de diámetro aproximadamente, se abre
dos días consecutivos; los pétalos presentan en la cara
adaxial dos pequeños nectarios;las flores son protoginas,
el primer día los 3 pistilos están separados, y los
estambres son cortos e indehiscentes. |
Cabomba caroliniana,
flor |
|

|
|
Antesis de Cabomba: 1º
y 2º día |
El 2º día los pistilos se
juntan en el centro, y están rodeados por los estambres
que se han alargado, de manera que cada antera queda por
encima de un nectario.
Cuando las moscas visitan las flores en su segundo
día de antesis, al libar el néctar su cabeza toma
contacto con la antera y se carga de polen.
Al visitar flores en su primer día de antesis, al
buscar el néctar, la cabeza deposita el polen sobre los
estigmas. |

|
|
Moscas visitando
flores en su 2º día
|
 |
| Esquemas y fotos de
Schneider & Jeter 1982 |
| Otras flores polinizadas por
dípteros tienen olor desagradable, a carroña, que
atrae
a las moscas necrófagas, como por ejemplo las flores de Rafflesia
arnoldii (Rafflesiaceae), planta del sudeste de
Asia, la flor más grande conocida. |
Flor de Rafflesia
arnoldi (casi un metro de diámetro) |
|

|
|
Imagen
de ??? |
Algunas especies polinizadas por dípteros presentan flores
trampa con
flores unisexuales protoginas. La
inflorescencia de Arum maculatum es un espádice, está
protegida
por la espata;
en la parte basal presenta flores femeninas, luego
están las flores masculinas fértiles, y luego flores estériles
(flores neutras) con una gruesa cerda orientada hacia abajo;
la
porción apical constituye un osmóforo que produce olor
fétido. La espata está dilatada en la base formando una urna,
estrechada a la altura de las flores estériles y ampliamente
abierta arriba formando el dispositivo de reclamo. La epidermis
interna es muy lisa, con gotitas de aceite secretadas por
papilas. Las moscas, atraídas por el olor emitido por el
osmóforo, resbalan y caen en la urna, de la que no
pueden salir por las cerdas de las flores neutras. A la noche
las flores masculinas fértiles liberan el polen que espolvorea
los insectos. Al día siguiente se marchitan las flores
masculinas estériles y los insectos cargados de polen pueden
salir y se precipitan generalmente en otra flor en la que
polinizan las flores femeninas.
Arum maculatum,
polinización miófila
|
|
Planta con
inflorescencia |
Inflorescencia: espata
cortada longitudinalmente en la base |
Detalle de la porción
florífera |
 |
 |
 |
|
Imagenes
de http://www.rz.uni-karlsruhe.de
|
Imagen de
http://bib18.ulb.ac.be |
Himenópteros (abejas
y avispas): flores melitófilas.
Estos insectos tienen aparato bucal suctor, buscan néctar accesible,
porque tienen órganos bucales cortos. Las flores nectaríferas tienen
un costo energético menor para la planta, por que el néctar constituye un ahorro de
polen.
Las flores melitófilas atraen a las abejas por una combinación de forma, olor y
color. Las corolas de estas flores son generalmente
amariposadas, labiadas o en fauce, con superficies para que la
abeja se pose, con guías (manchas o líneas coloreadas) que
señalan dónde se encuentra el néctar. Las flores producen sustancias aromáticas
en los osmóforos, que se
encuentran en la corola (Citrus), corona (Narcissus) u otros
órganos florales.
| Las abejas perciben el amarillo, el azul, el blanco;
no
perciben en cambio el rojo puro. También perciben los
rayos ultravioleta, no visibles por los ojos humanos, y
por esta razón, muchas flores que son amarillas a
nuestros ojos, a los ojos de las abejas se ven de un color
llamado "púrpura de abejas". |
"Púrpura de
abejas" en Oenothera |
|

|
|
Fotos de http://www.windows.ucar.edu |
Las abejas son capaces de memorizar sensaciones
(color y olor) de una visita favorable, y la repiten varios
días en flores de la misma especie: asocian nutrición con
atributos florales, y además pueden transmitir la información
a sus compañeras mediante el lenguaje por danza.
Las flores se abren gracias a la habilidad
de la recolectora; ciertas flores sólo pueden ser polinizadas
por animales con una determinada estructura corpórea. La flor
de Salvia
pratensis posee 2 estambres adnatos a la corola, en los
cuales el filamento es corto y el conectivo es
largo, con 2 brazos; el brazo largo lleva la teca fértil, y el
brazo corto una teca estéril convertida en una plaquita que se une
con la del otro estambre (Fig. C). Esta plaquita, en posición
normal (Fig. B) impide el libre acceso
al néctar. Al ser presionada por el polinizador (Fig. C), abejas o
abejorros del género Bombus, los extremos largos de los
conectivos descienden y sus tecan fértiles, unidas entre sí,
frotan el dorso del insecto depositando el polen (Fig. A). Las flores son
protándricas, así en flores viejas, el estilo se alarga, y estigma
receptivo ocupa el lugar
de las tecas fértiles (Fig. D), y cuando Bombus visita una de ellas deja
el polen en el estigma.
| Salvia
pratensis: polinización entomófila por abejas
Bombus |
 |
|
Dibujos de Strasburger
2004 |
En ciertas especies de plantas (
Ericaceae, Melastomataceae, Solanum spp., Cassia)
la recolección del polen es
vibrátil (buzz-pollination).
Es llevada a cabo por abejas o abejorros (Bombus).
Estos insectos se aferran a la flor y con rápidas contracciones de sus músculos para
vuelo indirecto producen el zumbido característico que hace vibrar las anteras
provocando la salida del polen. |
Bombus |
|

|
|
en.wikipedia.org |
| Las flores con polinización
vibrátil comparten algunas características: corola en forma de
bol o con pétalos reflexos, tamaño pequeño a mediano,
frecuentemente falta de néctar, anteras con dehiscencia
poricida, granos de polen pequeños o medianos, no grasos, de
superficie lisa. (Knudsen & Olesen 1993) . |
Cassia fistula,
lluvia de oro, flor e inflorescencia
|
 |
 |
Hay flores engañosas que
imitan la forma, pilosidad y olor de las hembras de ciertas
avispas o abejas. El caso mejor conocido es el de Ophrys
insectifera (D.46). (Proctor: 206-208, fig. 7.20), orquídea
del sur de Europa; es visitada únicamente por dos especies de
avispas del género Argogorytes. Los machos nacen en
primavera varias semanas antes que las hembras, y en sus primeros
vuelos son atraídos por la fragancia de las flores de Ophrys,
similar a las feromonas secretadas por las hembras (Proctor 206).
Además, el labelo presenta forma, color y textura similar a las
hembras. El proceso se designa como pseudo-copulación porque las
avispas macho intentan aparearse con la flor, y al hacerlo, entran
en contacto con la antera, trasladando los polinios de una flor a
otra en intentos sucesivos.
Ophrys
insectifera, planta y flor |
Argogorytes:
pseudocopulación |
 |
 |
 |
| http://linnaeus.nrm.se/ |
http://pdubois.free.fr/orchidees/ |
Foto de
http://bib18.ulb.ac.be |
Ficus carica, Higuera: el higo de Esmirna,
el higo comercial más importante, presenta sólo flores
femeninas longistilas. Para producir frutos necesita la
polinización cruzada con la higuera silvestre o cabrahigo, con
flores masculinas y femeninas. El polinizador es una avispa
diminuta, Blastophaga psenes. El cabrahigo tiene flores
masculinas fértiles, flores femeninas fértiles longistilas y
flores femeninas estériles brevistilas donde las avispas ponen
sus huevos, pues su ovipositor es tan largo como el estigma. En
primavera nacen las avispas, se aparean y las hembras fecundadas
salen cargadas de polen. Entran en los siconos de la higuera de
Esmirna; como éstos tienen solo flores longistilas, no
pueden depositar allí sus huevos, pero las polinizan.
|
Ficus carica,
higuera: sicono y tipos de flores |
Blastophaga |
 |
 |
| Dibujos de
Strasburger 2004 |
Proctor & Yeo, 1996 |
También existen otras recompensas, las llamadas
"flores de aceite" ofrecen aceites o cuerpos grasos secretados o
almacenados en glándulas especiales llamadas elaióforos. Las
flores de la familia Malpighiaceae, y de varias especies de orquídeas
presentan esta característica, que atrae a determinados grupos de
himenópteros (abejas Antophoridae).
Nierembergia hippomanica, chuscho (Solanaceae) tiene flores
con corola hipocrateriforme, con tubo largo y delgado y limbo
amplio, 5-lobado. Presenta pelos secretores en la parte
basal del limbo corolino y sobre la porción basal de los
estambres. Sus polinizadores más efectivos son dos especies
de Tapinotapsis, que recogen aceites de los pelos
glandulares, juntan polen, duermen y copulan en la flor. Las
abejas polinizan la flor al juntar polen, se posan sobre las
piezas fértiles, primero el abdomen toma contacto con el estigma
(Fig. B); luego al girar hacia abajo para poder recoger polen con
las patas traseras, el abdomen toma contacto con las anteras,
recogiendo polen que luego depositará en el estigma de otra flor
(Fig. C).
| Flores de Nierembergia |
Tapinotapsis en flor de Nierembergia |
Polinización de Nierembergia |
 |

|
 |
|
Dibujos de Cocucci 1984 y
1991 |
Lepidópteros (mariposas)
Las mariposas tienen aparato bucal suctor,
presentan espiritrompas muy largas que cuando no están en uso,
están enroscadas. Polinizan flores nectaríferas con el néctar
hundido en espolones o tubos largos. Se distinguen dos tipos:
1) Las mariposas diurnas visitan flores cuya
antesis tiene lugar durante el día (flores Psicófilas).
Son flores erectas, fragantes, coloridas, pueden ser rojas.
Frecuentemente las flores son tubulosas, o con el néctar profundamente escondido,
o con espolones
largos (Dianthus, Nicotiana tabacum), pero también son
visitadas por mariposas otras flores o inflorescencias
nectaríferas, como las de Eryngium.
|
Flores rojas,
tubulosas, psicófilas |
Nicotiana tabacum,
tabaco |
Eryngium
mesopotamicum, cardo |
 |

|
 |
| foto
de ??? |
foto
de ??? |
|
2) Las mariposas nocturnas visitan flores
estrechamente tubulosas, con antesis al anochecer, muy
perfumadas, horizontales o péndulas, blanquecinas o de colores
claros, pues
necesitan muy poca luz para apreciar los colores, (Lillium, Nicotiana, Lonicera, Cestrum
parqui).
Entre los lepidópteros nocturnos hay dos grupos: los esfíngidos,
mariposas grandes, robustas (flores Esfingófilas) y
las polillas (flores
Falenófilas) |
Nicotiana alata, flor
esfingófila |
|

|
|
Tegeticula yucasella es el nombre científico de la polilla que poliniza Yucca, deposita sus
huevos en el ovario. Durante el desarrollo del fruto, las larvas comen parte de los óvulos. |
Flor de Yucca
visitada por Tegeticula yucasella |
 |
|
Foto
de ??? |
Flores Ornitófilas.
En América los pájaros que se alimentan de néctar son los picaflores o
colibríes. Las aves tienen sensibilidad visual
similar al hombre, destacan el rojo. Visitan flores o inflorescencias
grandes, a menudo tubulosas o cóncavas, también flores o
inflorescencias en brocha. Los colores más frecuentes en las
flores visitadas por las aves son : rojo, anaranjado y amarillo.
Las flores a menudo son péndulas, sin superficies para posarse el ave. Las
flores presentan guías o señales de néctar, manchas o líneas
de la corola que llevan a los pájaros hacia los nectarios. El néctar
es fluído, abundante, escondido profundamente, lo absorben
con su lengua tubulosa o en pincel. El polen se adhiere al pico
o a otras partes de la cabeza. Las flores no tienen olor, ya que
los pájaros
tienen el sentido del olfato mal desarrollado. Las flores
ornitófilas son propias de
regiones tropicales: Hibiscus, Tropaeolum, Fuchsia, Euphorbia
pulcherrima, Erythrina cristagalli.
|
Flores ornitófilas:
colibríes libando el néctar en vuelo |
 |

|
 |
|
Fotos
de ??? |
Flores Quiropterófilas (murciélagos, con
lengua larga). Son típicas de los trópicos. Se trata de flores robustas, fasciculiformes o
cóncavas, con garganta ancha, de antesis nocturna, de colores obscuros,
fuerte olor a fruta o material fermentado, con gran cantidad de
néctar y polen. Musa, Agave, Adansonia (baobab, Bombacaceae),
algunas Bignoniáceas, Cactaceae en candelabro de flores
nocturnas.
Otros animales
Pueden participar en la polinización algunos marsupiales, pequeños
mamíferos y roedores que utilizan el néctar como alimento, pero desempeñan un papel secundario.
|
Flores
quiropterófilas: Passiflora y una Cactaceae |
Flor de Protea
polinizada por un ratón |
 |
 |
 |
| Fotos
de ??? |
|
www.naturalvisions.co.uk/ |
|
|