|
|
Major Groups of Prokaryotes© 1997 Kenneth Todar University of Wisconsin-Madison Department of Bacteriology  Major Groups of ProkaryotesThe prokaryotes consist of millions of genetically-distinct unicellular organisms. What they lack in structural diversity, so well-known among eukaryotes (including the protista), they make up for in their physiological diversity. It is often a particular physiological trait that unifies and distinguishes a particular group of prokaryotes. In Bergey's Manual (Reference 2) the groups of prokaryotes are formed based on easily-observed characteristics such as Gram stain, morphology (rods, cocci, etc), motility, structural features (e.g. spores, filaments, sheaths, appendages, etc.), and on distinguishing physiological features (e.g. anoxygenic photosynthesis, methanogenesis, lithotrophy, etc.). Nowadays, of course, the politically correct way to group organisms, especially prokaryotes, is on a genetic basis, i.e., by comparison of the nucleotide sequences of the small subunit ribosomal RNA that is contained in all cellular organisms. The ensuing description of prokaryotes may rely on various of these cellular traits in defining "groups", but herein the prokaryotes are placed under trivial headings based on common structural, biochemical or ecological properties. This does not imply close genetic relatedness among all members in a group. Sometimes, all of the members of a group share a close genetic relatedness; in other cases, members of a group are genetically-unrelated, even to an extent that is greater than exists among all members of the Eukaryotic domain. Also herein, some prokaryotes are in more than one group, and some groups consist of both Archaea and Bacteria. ArchaeaOn the basis of ssrRNA analysis, the Archaea consist of three phylogenetically-distinct groups: Crenarchaeota, Euryarchaeota and Korarchaeota. However, for the Korarchaeota, only their nucleic acids have been detected, and no organisms have been isolated or cultured. Based on their physiology, the Archaea can be organized into three types: methanogens (prokaryotes that produce methane); extreme halophiles (prokaryotes that live at very high concentrations of salt (NaCl); and extreme (hyper) thermophiles (prokaryotes that live at very high temperatures). Besides the unifying archaeal features that distinguish them from Bacteria (i.e., no murein in cell wall, ester-linked membrane lipids, etc.), these prokaryotes exhibit unique structural or biochemical attributes which adapt them to their particular habitats. The Crenarchaeota consists mainly of hyperthermophilic sulfur-dependent prokaryotes and the Euryarchaeota contains the methanogens and extreme halophiles. ssrRNAs of the Korarchaeota have been obtained from hyperthermophilic environments similar to those inhabitated by Crenarchaeota. 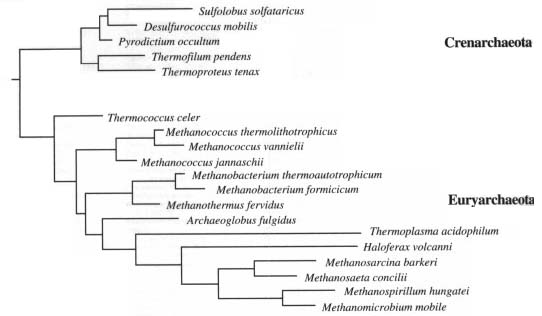 Methanogens are obligate anaerobes that will not tolerate even brief exposure to air (O2). Anaerobic environments are plentiful, however, and include marine and fresh-water sediments, bogs and deep soils, intestinal tracts of animals, and sewage treatment facilities. Methanogens have an incredible type of metabolism that can use H2 as an energy source and CO2 as a carbon source for growth. In the process of making cell material from H2 and CO2, the methanogens produce methane (CH4) in a unique energy-generating process. The end product (methane gas) accumulates in their environment. Methanogen metabolism created most the natural gas (fossil fuel) reserves that are tapped as energy sources for domestic or industrial use. Methanogens are normal inhabitants of the rumen (fore-stomach) of cows and other ruminant animals. A cow belches about 50 liters of methane a day during the process of eructation (chewing the cud). Methane is a significant greenhouse gas and is accumulating in the atmosphere at an alarming rate. When rain forests are destroyed and replaced by cows, it is "double-hit" on the greenhouse: (1) less CO2 is taken up by removal of the the autotrophic green plants; (2) additional CO2 and CH4 are produced as gases by the combined metabolism of the animal and methanogens. Methanogens represent a microbial system that can be exploited to produce energy from waste materials. Large amounts of methane are produced during industrial sewage treatment processes, but the gas is usually wasted rather than trapped for recycling. Figure 2. Methanococcus jannischii 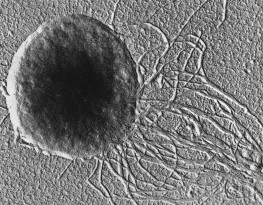
Methanococcus jannischii was originally isolated from a sample taken from a "white smoker" chimney at an oceanic depth of 2,600 meters on the East Pacific Rise. It can be grown in a mineral medium containing only H2 and CO2 as sources of energy and carbon for growth within a temperature range of 50 to 86 degrees. Cells are irregular cocci that are motile due to two bundles of polar flagella inserted near the same cellular pole. Extreme halophiles live in natural environments such as the Dead Sea, the Great Salt Lake, or evaporating ponds of seawater where the salt concentration is very high (as high as 5 molar or 25 percent NaCl). These prokaryotes require salt for growth and will not grow at low salt concentrations. Their cell walls, ribosomes, and enzymes are stabilized by Na+. Halobacterium halobium, the prevalent species in the Great Salt Lake, adapts to the high-salt environment by the development of "purple membrane", actually patches of light-harvesting pigment in the plasma membrane. The pigment is a type of rhodopsin called bacteriorhodopsin which reacts with light in a way that forms a proton gradient on the membrane allowing the synthesis of ATP. This is the only example in nature of non photosynthetic photophosphorylation. These organisms are heterotrophs that normally respire by aerobic means. The high concentration of NaCl in their environment limits the availability of O2 for respiration so they are able to supplement their ATP-producing capacity by converting light energy into ATP using bacteriorhodopsin. Figure 3. Halobacterium salinarium 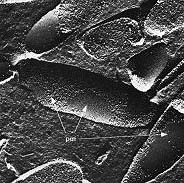
Halobacterium salinarium is an extreme halophile that grows at 4 to 5 M NaCl and does not grow below 3 M NaCl. This freeze etched preparation shows the surface structure of the cell membrane and reveals smooth patches of "purple membrane" (bacteriorhodopsin) embedded in the plasma membrane. The extreme thermophiles come from several distinct phylogenetic lines of Archaea. These organisms require a very high temperature (80 degrees to 105 degrees) for growth. Their membranes and enzymes are unusually stable at high temperatures. Most of these Archaea require elemental sulfur for growth. Some are anaerobes that use sulfur as an electron acceptor for respiration in place of oxygen. Some are lithotrophs that oxidize sulfur as an energy source. Sulfur-oxidizers grow at low pH (less than pH 2) because they acidify their own environment by oxidizing So (sulfur) to SO4 (sulfuric acid). These hyperthermophiles are inhabitants of hot, sulfur-rich environments usually associated with volcanism, such as hot springs, geysers and fumaroles in Yellowstone National Park, and thermal vents ("smokers") and cracks in the ocean floor. Sulfolobus was the first hyperthermophilic Archaean discovered by Thomas D. Brock of the University of Wisconsin in 1970. His discovery, along with that of Thermus aquaticus in Yellowstone National Park, launched the field of hyperthermophile biology. (Thermus aquaticus, source of the enzyme taq polymerase used in the polymerase chain reaction (PCR), is a Bacterium which has an optimum temperature for growth of 70 degrees.) Sulfolobus grows in sulfur-rich, hot acid springs at temperatures as high as 90 degrees and pH values as low as 1. Thermoplasma, also discovered by Brock, is a unique thermophile that is the sole representative of a distinct phylogenetic line of Archaea. Thermoplasma resembles the bacterial mycoplasmas in that it lacks a cell wall. Thermoplasma grows optimally at 55 degrees and pH 2. Interestingly, it has only been found in self-heating coal refuse piles, which are a man-made waste. Figure 4. Sulfolobus acidocaldarius 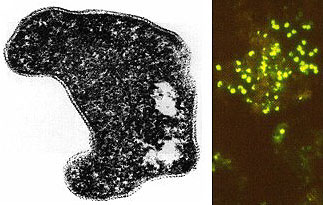
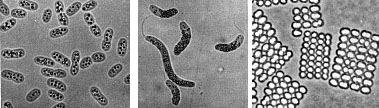
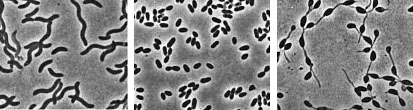
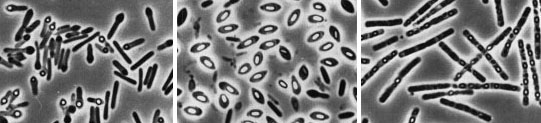
(T.D. Brock) Sulfolobusis an extreme thermophile that has been found in geothermally-heated acid springs, mud pots and surface soils with temperatures from 60 to 95 degrees C, and a pH of 1 to 5. Left: Electron micrograph of a thin section (X85,000). Under the electron microscope the organism appears as irregular spheres which are often lobed. Right: Fluorescent photomicrograph of cells attached to a sulfur crystal. Fimbrial-like appendages have been observed on the cells attached to solid surfaces such as sulfur crystals. Although the Archaea are often inhabitants of unusual or extreme environments, there may be corresponding species of Bacteria, and even eukaryotes, in these habitats as well. No bacterium can produce methane, but Bacteria are always found in anaerobic habitats in association with methanogens. With regard to acid tolerance, a bacterium, Thiobacillus, has been observed growing at pH near 0. An Alga, Cyanidium, has also been found growing near pH 0. In superheated environments (greater than100 degrees), Archaea seem to have an exclusive hold, but Bacteria have been isolated from boiling hot springs in Yellowstone National Park and other parts of the world. No bacterium grows at the highest salt concentration which supports the growth of the halobacteria, but osmophilic yeasts and fungi can grow at correspondingly low water actvities where sugar is the solute in high concentration. BACTERIAPhylogenetic analysis of the Bacteria has demonstrated that eleven distinct groups exist but most groups consist of members that are phenotypically and physiologically unrelated. Below we discuss the major groups of Bacteria based on morphology, physiology, or ecology, and often use informal, but familiar, terms to identify them.  Photosynthetic purple and green bacteria. These bacteria conduct anoxygenic photosynthesis, also called bacterial photosynthesis. Bacterial photosynthesis differs from plant-type (oxygenic) photosynthesis in several ways. Bacterial photosynthesis does not produce O2; in fact, it only occurs under anaerobic conditions. Bacterial photosynthesis utilizes chlorophyll other than chlorophyll a, and only one photosystem, photosystem I. The electron donor for bacterial photosynthesis is never H2O but may be H2, H2S of So, or certain organic compounds. The light-absorbing pigments of the purple and green bacteria consist of bacterial chlorophylls and carotenoids. Phycobilins, characteristic of the cyanobacteria, are not found. Many purple and green sulfur bacteria store elemental sulfur as a reserve material that can be further oxidized to SO4 as a photosynthetic electron donor. The purple and green sulfur bacteria use H2S during photosynthesis in the same manner that cyanobacteria or algae or plants use H2O as an electron donor for autotrophic CO2 reduction (the "dark reaction" of photosynthesis). They can also use organic compounds as electron donors for photosynthesis. For example, Rhodobacter can use light as an energy source while oxidizing succinate or butyrate in order to obtain electrons for CO2 fixation.  Figure 8. Photosynthetic prokaryotes growing in a hot spring run-off channel (T.D. Brock). The white area of the channel is too hot for photosynthetic life, but as the water cools along a gradient, the colored phototrophic bacteria colonize and ultimately construct the colored microbial mats composed of a consortium of photosynthetic microorganisms. Cyanobacteria. The cyanobacteria deserve special emphasis because of their great ecological importance in the global carbon, oxygen and nitrogen cycles, as well as their evolutionary significance in relationship to plants. Photosynthetic cyanobacteria have chlorophyll a and carotenoids in addition to some unusual accessory pigments named phycobilins. The blue pigment, phycocyanin and the red one, phycoerythrin, absorb wavelengths of light for photosynthesis that are missed by chlorophyll and the carotenoids. Within the cytoplasm of cyanobacteria are numerous layers of membranes, often parallel to one another. These membranes are photosynthetic thylakoids that resemble those found in chloroplasts, which, in fact, correspond in size to the entire cyanobacterial cell. The main storage product of the cyanobacteria is glycogen, and glycogen inclusions may be seen in the cytoplasm of the cells. Cyanobacteria are thought to have given rise to eukaryotic chloroplasts during the evolutionary events of endosymbiosis. In biochemical detail, cyanobacteria are especially similar to the chloroplasts of red algae (Rhodophyta). Most cyanobacteria have a mucilaginous sheath, or coating, which is often deeply pigmented, particularly in species that occur in terrestrial habitats. The colors of the sheaths in different species include light gold, yellow, brown, red, green, blue, violet, and blue-black. It is these pigments that impart color to individual cells and colonies as well as to "blooms" of cyanobacteria in aquatic environments  Figure 9. Some common cyanobacteria L to R: Oscillatoria, a filamentous species common in fresh water and hot springs; Nostoc, a sheathed communal species; Anabaena, a nitrogen fixing species. The small cell with an opaque surface (third from right) in the anabaena filament is a heterocyst, a specialized cell for nitrogen fixation. The large bright cell in the filament is a type of spore called an akinete; Synechococcus, a unicelluar species in marine habitats and hot springs. Synechococcus is among the most important photosynthetic bacteria in the marine environment, estimated to account for about 25 percent of the primary production that occurs in typical marine habitats. Although thousands of cyanobacteria have been observed, only about 200 species have been identified as distinct, free-living, nonsymbiotic prokaryotes. Relative to other oxygenic phototrophs, cyanobacteria often grow under fairly extreme environmental conditions such as high temperature and salinity . They are the only oxygenic phototrophs present in many hot springs of the Yellowstone ecosystem, and in frigid lakes and oceans of Antarctica, they form luxuriant mats 2 to 4 centimeters thick in water beneath more than five meters of permanent ice. However, cyanobacteria are absent in acidic waters where their eukaryotic counterparts, the algae, may be abundant. Layered chalk deposits called stromatolites, which exhibit a continuous geologic record covering 2.7 billion years, are produced when colonies of cyanobacteria bind calcium-rich sediments. Today, stromatolites are formed in only a few places, such as shallow pools in hot dry climates. The abundance of cyanobacteria in the fossil record is evidence of the early development of the cyanobacteria and their important role in elevating the level of free oxygen in the atmosphere of the early Earth. Cyanobacteria often form filaments and may grow in large masses or "tufts" one meter or more in length. Some are unicellular, a few form branched filaments, and a few form irregular plates or irregular colonies. Cyanobacterial cells usually divide by binary fission, and the resulting progeny cells may separate to form new colonies. In addition, filaments may break into fragments, called hormogonia, which separate and develop into new colonies. As in other filamentous or colonial bacteria, the cells of cyanobacteria may joined by their walls or by mucilaginous sheaths, but each cell is an independent unit of life. As true Bacteria, cyanobacteria contain peptidoglycan or murein in their cell walls. Most cyanobacteria have a Gram-negative type cell wall that consists of an outer membrane component, even though they show a distant phylogenetic relationship with Gram-positive bacteria. Some of the filamentous cyanobacteria are motile by means gliding or rotating around a longitudinal axis. Short segments (hormogonia) may break off from a cyanobacterial colony and glide away from their parent colony at rates as rapid as 10 micrometers per second. The mechanism for this movement is unexplained but may be connected to the extrusion of slime (mucilage) through small pores in their cell wall, together with contractile waves in one of the surface layers of the wall. Cyanobacteria are found in most aerobic environments where water and light are available for growth. Mainly they live in fresh water and marine habitats. Those inhabiting the surface layers of water are part of a complex microbial community called plankton. Planktonic cyanobacteria usually contain cytoplasmic inclusions called gas vesicles which are hollow protein structures filled with various gases. The vesicles can be inflated or deflated with gases allowing the organisms maintain buoyancy and to float at certain levels in the water. Thus, the cyanobacteria can regulate their position in the water column to meet their optimal needs for photosynthesis, oxygen, and light-shielding. When numerous cyanobacteria become unable to regulate their gas vesicles properly (for example, because of extreme fluctuations of temperature or oxygen supply), they may float to the surface of a body of water and form visible "blooms". A planktonic species related to Oscillatoria gives rise to the redness (and the name) of the Red Sea. The cyanobacteria have very few harmful effects on plants or animals. They may be a nuisance if they bloom in large numbers and then die and decay in bodies of fresh water that are used for drinking or recreational purposes. Many cyanobacteria are responsible for the earthy odors and flavors of fresh waters, including drinking waters, due to the production of compounds called geosmins. Some cyanobacteria that form blooms secrete poisonous substances that are toxic for animals that ingest large amounts of the contaminated water. Many marine cyanobacteria occur in limestone (calcium carbonate) or lime-rich substrates, such as coral algae and the shells of mullosks. Some fresh water species, particularly those that grow in hot springs, often deposit thick layers of lime in their colonies. Some cyanobacteria can fix nitrogen. In filamentous cyanobacteria, nitrogen fixation often occurs in heterocysts, which are specialized, enlarged cells, usually distributed along the length of a filament or at the end of a filament. Heterocysts have intercellular connections to adjacent vegetative cells, and there is continuous movement of the products of nitrogen fixation moving from heterocysts to vegetative cells, and the products of photosynthesis moving from vegetative cells to heterocysts. Heterocysts are low in phycobilin pigments and have only photosystem I. They lack the oxygen-evolving photosystem II. Furthermore, they are surrounded in a thickened, specialized glycolipid cell wall that slows the rate of diffusion of O2 into the cell. Any O2 that diffuses into the heterocyst is rapidly reduced by hydrogen, a byproduct of N2 fixation, or is expelled through the wall of the heterocyst. The process of nitrogen fixation, specifically the enzyme nitrogenase, only functions in anaerobic conditions so the organism must maintain these oxygen-free compartments in order for N2 fixation to occur. In addition to the heterocysts, some cyanobacteria form resistant spores called akinetes enlarged cells around which thickened outer walls develop. Akinetes are resistant to heat, freezing and drought (desiccation) and thus allow the cyanobacteria to survive unfavorable environmental conditions. The are functionally analogous to bacterial endospores, but they bear little resemblance and lack the extraordinary resistance properties of endospores. A few cyanobacteria are symbionts of liverworts, ferns, cycads, flagellated protozoa, and algae, sometimes occurring as endosymbionts of the eukaryotic cells. In the case of the water fern Azolla, the cyanobacterial endophyte (a species of Anabaena) fixes nitrogen that becomes available to the plant . In addition, it is often the case that the photosynthetic partners of lichens are cyanobacteria. The planktonic cyanobacteria fix an enormous amount of CO2 during photosynthesis, and as "primary producers" they and are the basis of the food chain in marine environments. Their type of photosynthesis, which utilizes photosystem II, generates a substantial amount of oxygen present in the earth's atmosphere. Since many cyanobacteria can fix N2 under certain conditions, they are one of the most significant free-living nitrogen-fixing prokaryotes. Cyanobacteria carried out plant-type (oxygenic) photosynthesis for at least a billion and a half years before the emergence of plants, and cyanobacteria are believed to be the evolutionary forerunners of modern-day plant and algal chloroplasts. A group of phototrophic prokaryotes, called prochlorophytes contain chlorophyll a and b but do not contain phycobilins. Prochlorophytes, therefore, resemble both cyanobacteria (because they are prokaryotic and contain chlorophyll a) and the plant chloroplast (because they contain chlorophyll b instead of phycobilins). Prochloron, the first prochlorophyte discovered, is phenotypically very similar to certain plant chloroplasts and is the leading candidate for the type of bacterium that might have undergone endosymbiotic events that led to the development of the plant chloroplast. Spirochetes are a phylogenetically distinct group of Bacteria which have a unique cell morphology and mode of motility. Spirochetes are very thin, flexible, spiral-shaped prokaryotes that move by means of structures called axial filaments or endoflagella. The flagellar filaments are contained within a sheath between the cell wall peptidoglycan and an outer membrane. The filaments flex or rotate within their sheath which causes the cells to bend, flex and rotate during movement. Most spirochetes are free living (in muds and sediments), or live in associations with animals (e.g. in the oral cavity or GI tract). A few are pathogens of animals (e.g. leptospirosis in dogs, Syphilis in humans and Lyme Disease in dogs and humans).  Figure 10. Spirochetes: A. Cross section of a spirochete showing the location of endoflagella between the inner membrane and outer sheath; B. Borrelia burgdorferi, the agent of Lyme disease; C. Treponema pallidum, the spirochete that causes syphilis. Spirilla are Gram-negative aerobic heterotrophic bacteria with a helical or spiral shape. Their metabolism is respiratory and never fermentative. Unlike spirochetes, they have a rigid cell wall and are motile by means of ordinary polar flagella. Spirilla are inhabitants of microaerophilic aquatic environments. Most spirilla require or prefer that oxygen in their environment be present in an amount that is well below atmospheric concentration. Spirillum and Aquaspirillum are inhabitants of fresh water. They frequently contain magnetosomes and exhibit the property of magnetotaxis (movement in relationship to the magnetic field of the earth). Oceanospirillum lives in marine habitats and is able to grow at NaCl concentrations as high as 9 percent. Azospirillum is a nitrogen-fixing bacterium that enters into a mutualistic symbiosis with certain tropical grasses and grain crops. Spirilla are thought to play a significant role in recycling of organic matter, particularly in aquatic environments. Related to the spirilla is the small vibrioid bacterium, Bdellovibrio, which is a parasite of other Gram-negative bacteria, including E. coli. It preys on other bacteria by entering into the periplasmic space and obtaining nutrients from the cytoplasm of its host cell while undergoing an odd type of reproductive cycle. Two pathogens of humans are found among the spirilla. Campylobacter jejuni is an important cause of bacterial diarrhea, especially in children. The bacterium is transmitted via contaminated food, usually undercooked poultry or shellfish, or untreated drinking water. Helicobacter pylori is able to colonize the gastric mucosal cells of humans, i.e., the lining of the stomach, and it has been fairly well established as the cause of peptic ulcers. The Myxobacteria are a group of gliding bacteria that aggregate together to form a multicellular fruiting body in which development and spore formation takes place. They exhibit the most complex behavioral patterns and life cycles of all known prokaryotes. Myxobacteria are inhabitants of the soil. They have a eukaryotic counterpart in nature in the Myxomycetes, or slime molds, and the two types of organisms are an example of parallel or convergent evolution, having adopted similar life styles in the soil environment. The vegetative cells of myxobacteria are typical Gram-negative rods that glide across a substrate such as a decaying leaf or piece of animal dung, or colonies of other bacteria. They obtain nutrients from the substrate as they glide across it and they secrete a slime track which other myxobacterial cells preferentially follow. If their nutrients become exhausted, the cells signal to one another to aggregate and form a swarm of myxobacteria which eventually differentiate into a multicellular fruiting body that contains myxospores, a type of dormant cell descended from a differentiated vegetative cell. In the case of Stigmatella, the myxospores are packed into secondary structures called cysts, which develop at the tips of the fruiting body (Figure 11). The bright-colored fruiting bodies of myxobacteria, containing millions of cells and spores, can often be seen with the unaided eye on dung pellets and decaying vegetation in the soil. 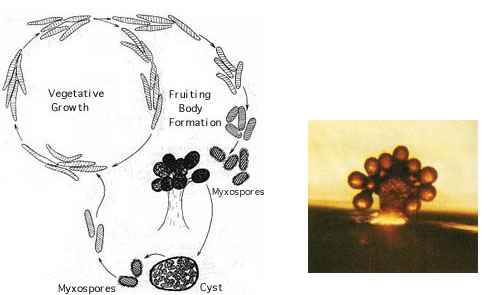 Figure 11. Stigmatella aurantiaca, a fruiting myxobacterium: L. Life Cycle R. Fruiting Body. Lithotrophs. Lithotrophy, a type of metabolism that requires inorganic compounds as sources of energy, is established in both the Archaea and the Bacteria. The methanogens utilize H2 as an energy source, and many extreme thermophiles use H2S or elemental sulfur as a source of energy for growth. Lithotrophic Bacteria are typically Gram-negative species that utilize inorganic substrates including H2, NH3, NO2, H2S, S, Fe++, and CO. Ecologically, the most important lithotrophic Bacteria are the nitrifying bacteria, Nitrosomonas and Nitrobacter that together convert NH3 to NO2, and NO2 to NO3, and the colorless sulfur bacteria such as Thiobacillus that oxidizes H2S to S and S to SO4. Most lithotrophic bacteria are autotrophs, and, in some cases, they may play an important role in primary production of organic material in nature. Lithotrophic metabolism does not extend to eukaryotes (unless a nucleated cell harbors lithotrophic endosymbiotic bacteria), and these bacteria are important in the biogeochemical cycles of the elements.  Figure 12. Lithotroph Habitats. A. Stream in Northern Wisconsin near Hayward is a good source of iron bacteria. B. Bacteriologist J.C. Ensign of the University of Wisconsin observing growth of iron bacteria in a run-off channel from the Chocolate Pots along the Gibbon River, in Yellowstone National Park. C. An acid hot spring at the Norris Geyser Basin in Yellowstone is rich in iron and sulfur. D. A black smoker chimney in the deep sea emits iron sulfides at very high temperatures (270 to 380 degrees C). Pseudomonads and their relatives "Pseudomonads" is a term for bacteria which morphologically and physiologically resemble members of the genus Pseudomonas, a very diverse group of Gram-negative rods with a strictly-respiratory mode of metabolism. Usually the term is reserved for members of the genera Pseudomonas and Xanthomonas, but many other related bacteria share the definitive characterictics of pseudomonads, i.e., Gram-negative bacteria which typically live by aerobic (as opposed to facultative), means. In Bergey's Manual, these bacteria are unified as Gram-negative aerobic rods and cocci. In Woese's Universal Phylogenetic Tree the genera are scattered about among the Purple Bacteria, with some being close relatives of the Enterobacteriaceae. In fact, the morphology and habitat of many pseudomonads sufficiently overlaps with the enterics (below) that microbiologists must quickly learn how to differentiate these two types of Gram-negative motile rods. Pseudomonads move by polar flagella; enterics such as E. coli swim by means of peritrichous flagella. Enterics ferment sugars such as glucose; pseudomonads generally do not ferment sugars. And most pseudomonads have an unusual cytochrome in their respiratory electron transport chain that can be detected in colonies by a colorimetric test called the oxidase test. Pseudomonads are oxidase- positive. 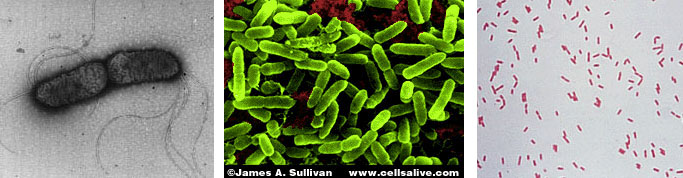 Figure 13. Profile of a Pseudomonad: Gram-negative rods motile by polar flagella. A. Electron micrograph, negative stain. B. Scanning electron micrograph. C. Gram stain. Most pseudomonads are free-living organisms in soil and water; they play an important role in decomposition, biodegradation, and the C and N cycles. The phrase "no naturally-occurring organic compound cannot be degraded by some microorganism" must have been coined to apply to members of the genus Pseudomonas, known for their ability to degrade hundreds of different organic compounds including insecticides, pesticides, herbicides, plastics, petroleum substances, hydrocarbons and other of the most refractory molecules in nature. However, they are usually unable to degrade biopolymers in their environment, such as cellulose and lignen, and their role in anaerobic decomposition is minimal. There are about 150 species of Pseudomonas, but, especially among the plant pathogens, there are many strains and biovars among the species. These bacteria are frequently found as part of the normal flora of plants, but they are one of the most important bacterial pathogens of plants, as well. Pseudomonas syringae and Xanthomonas species cause a wide variety of plant diseases as discussed below. One strain of Pseudomonas that lives on the surfaces of plants can act as an "ice nucleus" which causes ice formation and inflicts frost damage on plants at one or two degrees above the conventional freezing temperature of water (0 degrees C). One Pseudomonas species is an important pathogen of humans, Pseudomonas aeruginosa, the quintessential opportunistic pathogen, which is a leading cause of hospital-acquired infections. A few relatives of the pseudomonads are pathogens of animals, i.e., the agents of whooping cough, Bordetella pertussis, Legionaires' pneumonia (Legionella pneumophilia), gonorrhea (Neisseria gonorrhoeae), and bacterial meningitis (Neisseria meningitidis), but most of these bacteria have a normal existence apart from animals - in soil, water, and on the surfaces of plants. The pseudomonads have a lesser predilection for pathogenicity of animals than do the enteric bacteria. Among the interesting or important relatives of the pseudomonads are Rhizobiumand Bradyrhizobium, species that fix nitrogen in association with leguminous plants, and related Agrobacterium species that cause tumors ("galls") in plants. These bacteria are discussed later in this article because of their special relationships with plants. Relatives of the pseudomonads also include the methanotrophs that can oxidize methane and other one-carbon compounds, the azotobacters, which are very prevalent free-living (nonsymbiotic) nitrogen-fixing bacteria, and the acetobacters that are used in the manufacture of vinegar. Enterics. Enteric bacteria are Gram-negative rods with facultative anaerobic metabolism that live in the intestinal tracts of animals. This group consists of Escherichia coli and its relatives, the members of the family Enterobacteriaceae. Enteric bacteria are related phenotypically to several other genera of bacteria such as Pseudomonas and Alcaligenes, but are physiologically quite unrelated. Generally, a distinction can be made on the ability to ferment glucose; enteric bacteria all ferment glucose to acid end products while similar Gram-negative bacteria cannot ferment glucose. Because they are consistent members of the normal flora of humans, and because of their medical importance, an extremely large number of enteric bacteria have been isolated and characterized. Escherichia coli is, of course, the type species of the enterics. E. coli is such a regular inhabitant of the intestine of humans that it is used by public health authorities as an indicator of fecal pollution of drinking water supplies, swimming beaches, foods, etc. E. coli is the most studied of all organisms in biology because of its occurrence, and the ease and speed of growing the bacteria in the laboratory. It has been used in hundreds of thousands of experiments in cell biology, physiology, and genetics, and was among the first cells for which the entire chromosomal DNA base sequence was determined. In spite of the knowledge gained about the molecular biology and physiology of E. coli, surprisingly little is known about its ecology, for example why it consistently associates with humans, how it helps its host, how it harms its host, etc. A few strains of E. coli are pathogenic (one is notorious, strain 0157:H7, that keeps turning up in raw hamburger headed for a fast-food restaurants).Pathogenic strains of E. coli cause intestinal tract infections (usually acute and uncomplicated, except in the very young ) or uncomplicated urinary tract infections. 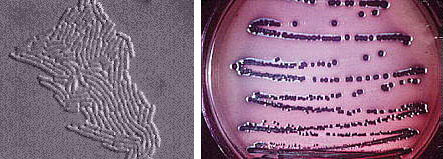 Figure 14. Left: Escherichia coli cells. Right: E. coli colonies on EMB Agar. The enteric group also includes some other intestinal pathogens of humans such as Shigella dysenteriae, cause of bacillary dysentery, and Salmonella typhimurium, cause of gastroenteritis. Salmonella typhi, which infects via the intestinal route, causes typhoid fever. Some bacteria that don't have an intestinal habitat resemble E. coli in enough ways to warrant inclusion in the enteric group. This includes Proteus, a common saprophyte of decaying organic matter, Yersinia pestis which causes bubonic plague, and Erwinia, an important pathogen of plants. Vibrios (which have a curved rod morphology or comma shape) are very common bacteria in aquatic environments. They have structural and metabolic properties that overlap with both the enterics and the pseudomonads. In Bergey's Manual, Vibrionaceae is a family on the level with Enterobacteriaceae. Vibrios are facultative like enterics, but they have polar flagella, are oxidase-positive, and dissimilate sugars in the same manner as the pseudomonads. In aquatic habitats they overlap with the Pseudomonadaceae in their ecology, although pseudomonads favor fresh water and vibrios prefer salt water. The genus Vibrio contains an important pathogen of humans, Vibrio cholerae, the cause of Asiatic cholera. Cholera is an intestinal disease with a pathology related to diarrheal diseases caused by the enteric bacteria. Five species of marine vibrios exhibit the property of bioluminescence, the ability to emit light of a blue-green color. These bacteria may be found as saprophytes on dead fish or as symbionts of living fish and invertebrates in marine environments. Some grow in special organs of the fish and emit light for the benefit of the fish (to attract prey, or as a mating signal) in return for a protected habitat and supply of nutrients. The reaction leading to light emission, catalyzed by the enzyme luciferase, has been found to be the same in all prokaryotes, and differs from light emission by eukaryotes such as the fire fly. Luciferase diverts electrons from the normal respiratory electron transport chain and causes formation of an excited peroxide that leads to emission of light. Nitrogen-fixing organisms. This is a diverse group of prokaryotes, reaching into phylogenetically distinct groups of Archaea and Bacteria. Members are unified only on the basis of their metabolic ability to "fix" nitrogen. Nitrogen fixation is the reduction of N2 (atmospheric nitrogen) to NH3 (ammonia). It is a complicated enzymatic process mediated by the enzyme nitrogenase. Nitrogenase is found only in prokaryotes and is second only to RUBP carboxylase (the enzyme responsible for CO2 fixation) as the most abundant enzyme on Earth. The conversion of nitrogen gas (which constitutes about 80 percent of the atmosphere) to ammonia introduces nitrogen into the biological nitrogen cycle. Living cells obtain their nitrogen in many forms, but usually from ammonia (NH3) or nitrates (NO3), and never from N2. Nitrogenase extracts N2 from the atmosphere and reduces it to NH3 in a reaction that requires substantial reducing power (electrons) and energy (ATP). The NH3 is immediately assimilated into amino acids and proteins by subsequent cellular reactions. Thus, nitrogen from the atmosphere is fixed into living (organic) material.
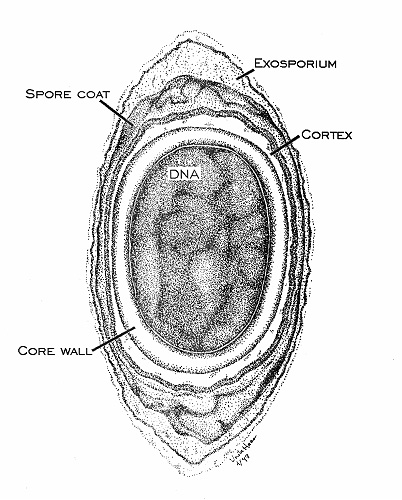 Figure 17. Anatomy of an endospore, cross section drawing by Viake Haas. Endospores differ from the vegetative cells that form them in a variety of ways. Several new surface layers develop outside the core (cell) wall, including the cortex and spore coat. The cytoplasm is dehydrated and contains only the cell genome and a few ribosomes and enzymes. The endospore is cryptobiotic (exhibits no signs of life) and is remarkably resistant to environmental stress such as heat (boiling), acid, irradiation, chemicals and disinfectants. Some endospores have remained dormant for 25 million years preserved in amber, only to be shaken back into life when extricated and introduced into a favorable environment. 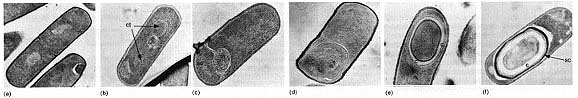
Figure 18. The sequential steps in the process of endospore formation in Bacillus subtilis. Some sporeformers are pathogens of animals, usually due to the production of powerful toxins. Bacillus anthracis causes anthrax, a disease of domestic animals (cattle, sheep, etc.) which may be transmitted to humans. Clostridium botulinum causes botulism, a form of food poisoning. Clostridium tetani is the agent of tetanus. 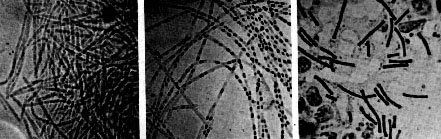 Figure 19. Robert Koch's original photomicrographs of Bacillus anthracis. In 1876, Koch established by careful microscopy that the bacterium was always present in the blood of animals that died of anthrax. He took a small amount of blood from such an animal and injected it into a healthy mouse, which subsequently became diseased and died. He took blood from that mouse and injected it into a another healthy mouse. After repeating this several times he was able to recover the original anthrax organism from the dead mouse, demonstrating for the first time that a specific bacterium is the cause of a specific disease. In so doing, he established Koch's Postulates, which still today supply the microbiological standard to demonstrate that a specific microbe is the cause of a specific disease. In association with the process of sporulation, some Bacillus species form a crystalline protein inclusion called parasporal crystals. The protein crystal and the spore (actually the spore coat) are toxic to lepidopteran insects (certain moths and caterpillars) if ingested. The crystals and spores of Bacillus thuringiensis are marketed as "Bt" a natural insecticide for use on garden or crop plants. Another species of Bacillus, B. cereus, produces an antibiotic that inhibits growth of Phytophthera, a fungus that attacks alfalfa seedling roots causing a "damping off" disease. The bacteria, growing in association with the roots of the seedlings, can protect the plant from disease. Also, apparently in association with the sporulation process, some Bacillus species produce clinically-useful antibiotics. Bacillus antibiotics such as polymyxin and bacitracin are usually polypeptide molecules that contain unusual amino acids. Actinomycetes and related bacteria are a large group of Gram-positive bacteria that usually grow by filament formation, or at least show a tendency towards branching and filament formation. Many of the organisms can form resting structures called spores, but they are not the same as endospores. Branched forms superficially resemble molds and are a striking example of convergent evolution of a prokaryote and a eukaryote together in the soil habitat. Actinomycetes such as Streptomyces have a world-wide distribution in soils. They are important in aerobic decomposition of organic compounds and have an important role in biodegradation and the carbon cycle. Products of their metabolism, called geosmins, impart a characteristic earthy odor to soils. Actinomycetes are the main producers of antibiotics in industrial settings, being the source of most tetracyclines, macrolides (e.g. erythromycin), and aminoglycosides (e.g. streptomycin, gentamicin, etc.). Two bacteria in this diverse group are important pathogens of humans: Mycobacterium tuberculosis is the cause of tuberculosis; Corynebacterium diphtheriae is the cause of diphtheria. Also, many nonpathogenic mycobacteria and corynebacteria live in associations with animals. 
Figure 20. Schematic diagrams illustrating mycelial growth and spore formation in several genera of actinomycetes. Rickettsias and chlamydiae are two unrelated groups of Bacteria that are obligate intracellular parasites of eukaryotic cells. Rickettsias cannot grow outside of a host cell because they have leaky membranes and are unable to obtain nutrients in an extracellular habitat. Chlamydiae are unable to produce ATP in amounts required to sustain metabolism outside of a host cell and are, in a sense, energy-parasites. Rickettsias occur in nature in the gut lining of arthropods (ticks, fleas, lice, etc.). They are transmitted to vertebrates by an arthropod bite and produce such diseases as typhus fever, Rocky Mountain Spotted Fever, Q fever and canine ehrlichiosis. Chlamydiae are tiny bacteria that infect birds and mammals. They may colonize and infect tissues of the eye and urogenital tract in humans. Chlamydia trachomatis causes several important diseases in humans: chlamydia, the most prevalent sexually transmitted disease in the U.S., trachoma, a leading cause of blindness worldwide, and lymphogranuloma venereum. 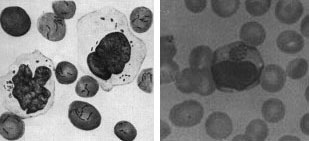
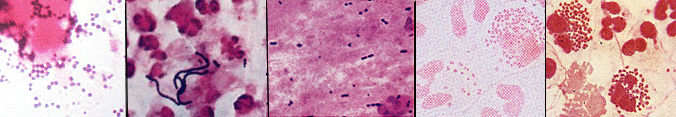
Figure 21. Mammalian cells infected with rickettsial organisms. L. Bartonella bacilliformis infection of human erythrocytes and blood monoyctyes. R. Ehrlichia canis infection of canine erythrocytes and blood monocytes. The distinct stained intracytoplasmic inclusion body in the monocyte is characteristic of the infection. Mycoplasmas are a group of bacteria that lack a cell wall. The cells are bounded by a single triple-layered membrane. They may be free-living in soil and sewage, parasitic inhabitants of the mouth and urinary tract of humans, or pathogens in animals and plants. In humans, Mycoplasma pneumoniae causes primary atypical pneumonia ("walking pneumonia"). Mycoplasmas include the smallest known cells, usually about 0.2 - 0.3 micrometers in diameter. Mycoplasmas correspondingly have the smallest known genome of any cell. Their DNA is thought to contain about 650 genes, which is about one-fifth the number found in E. coli and other common bacteria. Mycoplasmas can survive without a cell wall because their cytoplasmic membrane is more stable than that of other prokaryotes. In one group of mycoplasmas, the membrane contains sterols which seem to be responsible for the stability. Also, mycoplasmas tend to inhabit environments of high osmolarity wherein the risk of osmotic shock and lysis of the cells is minimized. Plant-pathogenic bacteria. Many economically-important diseases of plants are caused by members of the Bacteria. It is estimated that one-eighth of the crops worldwide are lost to diseases caused by bacteria, fungi or insects. Almost all kinds of plants can be affected by bacterial diseases, and many of these diseases can be extremely destructive. Almost all plant-pathogenic bacteria are Gram-negative bacilli, usually affiliated with the pseudomonads or enterics (above). The symptoms of bacterial disease in plants are described by a number of terms such as spots, blights, soft rots, wilts, and galls. Bacterial spots of various sizes on stems, leaves, flowers and fruits are usually caused by Pseudomonas or Xanthomonas species. Bacteria may cause spots by producing toxins that kill cells at the site of infection. Blights are caused by rapidly developing necrosis (dead, discolored areas) on stems, leaves and flowers. Fire blight in apples and pears, caused by Erwinia amylovora, can kill young trees within a single season. Bacterial soft rots occur most commonly in fleshy vegetables such as potatoes or onions or fleshy fruits such as tomatoes and eggplants. The most destructive soft rots are caused by Erwinia species that attack fruits and vegetables at the post-harvest stage. Bacterial vascular wilts mainly affect herbaceous plants. The bacteria invade the vessels of the xylem, where they multiply, interfering with the movement of water and inorganic nutrients and resulting in the wilting and the death of the plants. The bacteria commonly degrade portions of the vessel walls and can even cause the vessels to rupture. Once the walls have ruptured, the bacteria then spread to the adjacent parenchyma tissues, where they continue to multiply. In some bacterial wilts, the bacteria ooze to the surface of the stems or leaves through cracks formed over cavities filled with cellular debris, gums, and bacteria. More commonly, however, the bacteria do not reach the surface of the plant until the plant has been killed by the disease. Wilts of alfalfa and bean plants are cause by species of Clavibacter; bacterial wilt of cucurbits, such as squashes and watermelons, are cause by Erwinia tracheiphila; the black rot of crucifers such as cabbage is caused by Xanthomonas campestris. The most economically-important wilt of plants is caused by Pseudomonas solanacearum which affects 44 genera of plants, including such major crops as bananas, peanuts, tomatoes, potatoes, eggplants and tobacco. This disease occurs worldwide in tropical, subtropical, and warm temperate areas. Mycoplasmas (discussed above) have been identified in more than 200 plant species and associated with more than 50 plant diseases, many with symptoms of yellowing. Among these plant-pathogens are the spiroplasmas (genus Spiroplasma), which are pleomorphic, ovoid or spiral-shaped cells which are motile by means of a rotary or screw-like motion. Intracellular fibrils are thought to be responsible for their movement. The organisms have been isolated from the fluids of vascular plants and from the gut of insects that feed on these fluids. Some have been cultured on artificial media, including Spiroplasma citri, which is isolated from the leaves of citrus plants, where it causes citrus stubborn disease, and from corn plants suffering from corn stunt disease. A number of other mycoplasma-like organisms (sometimes called MLOs) have been detected in diseased plants by electron microscopy, which has been taken as evidence that these organisms may be more involved in plant disease than previously realized. The causative agent of a common plant disease, termed crown gall, is Agrobacterium tumefaciens. The disease is characterized by large galls or swellings that form on the plant at the site of infection, usually near the soil line. Crown gall is a problem in nurseries, affecting ornamental plants and fruit stock, and it may be a serious disease in grapes. Because of their role in the genetic engineering of plants, the molecular biology of these bacteria is intensively studied. References
Edited on Mar 25, 1998 by Kenneth Todar University of Wisconsin-Madison Department of Bacteriology |
|