Los protoplastos de las células vegetales al estar rodeados de pared celular tendrían dificultad para intercambiar material y para funcionar armónicamente, si no fuera por la existencia de comunicaciones
intercelulares:
Son conexiones citoplasmáticas que atraviesan la pared celular entre células contiguas. Al hallarse unidos entre sí los protoplastos de las células vivas por medio de
plasmodesmos, constituyen un simplasto único. El movimiento de sustancias a través de los plasmodesmos se denomina
transporte simplástico. Las paredes celulares, los lúmenes de las células muertas y los espacios intercelulares que rodean al simplasto formando también un continuo, se contraponen bajo el nombre de
apoplasto; el movimiento de sustancias en él se conoce como
transporte apoplástico.
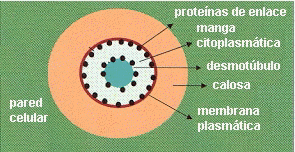
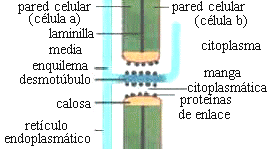
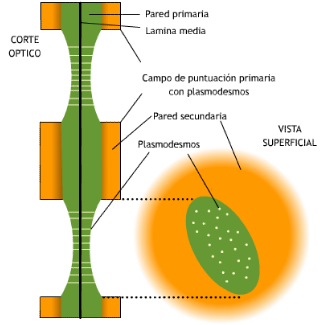
Si se observa un plasmodesmo en sección transversal con
MET (microscopio electrónico de transmisión), se ve una doble membrana: la externa es la membrana plasmática, rodeada por una delgada capa de calosa
, la interna corresponde al desmotúbulo, que es un túbulo del retículo endoplasmático, entre ambas hay una
manga citoplasmática . Los componentes de la cara interna de la biomembrana que forma el desmotúbulo se fusionan entre sí, de manera que el desmotúbulo no tiene lumen.
El
transporte entre célula y célula está limitado a la "manga citoplasmática" que rodea al desmotúbulo
(Fig. 7.15 y 7.16).
Fig.7.16, Diagrama
de plasmodesmo
entre dos células
Estudios con microscopía electrónica de alta resolución han demostrado que hay proteínas globulares de enlace ("linking proteins") incrustadas en la membrana plasmática que rodea al
plasmodesmo, y en la cara externa del desmotúbulo. Estas proteínas dividen la manga citoplasmática en microcanales que determinan el tamaño máximo de las moléculas que pueden desplazarse por difusión al mismo tiempo que establecen el tráfico selectivo de macromoléculas, que parece ocurrir por un proceso análogo al trasporte
núcleo-citoplasmático.
Los plasmodesmos primarios se forman durante la citocinesis al mismo tiempo que la pared celular. En casos especiales como los injertos, tejidos cicatriciales, interfase parásito-huésped, se forman plasmodesmos secundarios en lugares donde antes no existían o por modificación de plasmodesmos primarios. No se forman en paredes que al otro lado no tienen células vivas.
Cuando una célula muere engruesa rápidamente la cubierta de calosa en la célula vecina y se
oblitera el plasmodesmo. Ocasionalmente los plasmodesmos se ramifican a uno o ambos lados de la laminilla media; en ese caso se forma una cavidad.
Raramente se presentan esparcidos en las paredes primarias, a
veces en las que son bastante gruesas, como en las células del endosperma de
ciertas semillas como las de Diospyros.
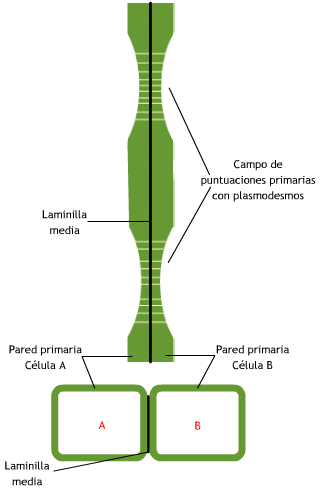
Comúnmente están agrupados en zonas adelgazadas, deprimidas de las
paredes primarias, constituyendo un campo primario de puntuación o
puntuación primordial. En el límite del campo primario de puntuación,
las microfibrillas se disponen paralelamente, formando un círculo u
óvalo (Fig. 7.17).
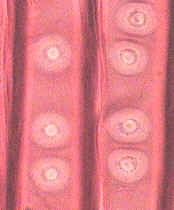
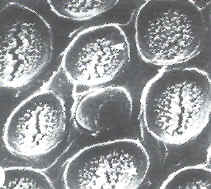
Los campos primarios de puntuaciones o de punteaduras, también llamadas puntuaciones primordiales, pueden ser tan abundantes que la
pared primaria en corte presenta aspecto arrosariado
(Fig. 7.18). Sucede especialmente cuando las células adyacentes cooperan estrechamente, como las células del mesófilo y la
vaina. En células fisiológicamente aisladas (estomas) el número de plasmodesmos es bajo.
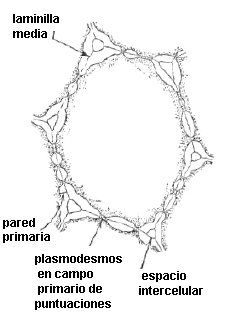
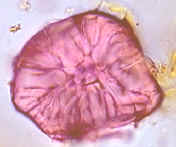
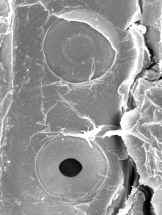
Fig.
7.17, Campo primario de puntuación en vista superficial (foto MEB,
microscopio electrónico de barrido)
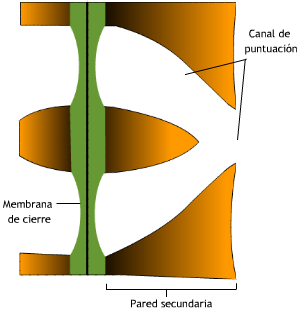
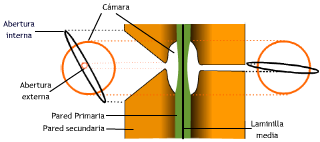
Las puntuaciones son discontinuidades en la deposición de la pared secundaria a nivel de un campo primario de puntuación, aunque también pueden diferenciarse en zonas donde no había campos primarios. Se distinguen dos tipos principales de puntuaciones:
Partes: la membrana de cierre o membrana alveolar formada por la laminilla media y pared primaria adelgazada; la
cavidad de la puntuación formada por la discontinuidad en la deposición de la pared secundaria, a veces tapizada por una capa
verrucosa.
Si la pared secundaria es muy gruesa, la cavidad forma el canal de la puntuación, que va desde el lumen hasta la membrana de cierre.
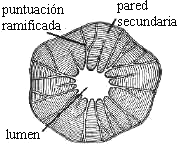
Como el tamaño del lumen se va reduciendo con el incremento en grosor de la pared, pueden fusionarse los canales de dos o más puntuaciones vecinas constituyendo entonces las llamadas
puntuaciones ramificadas (Fig. 7.20).
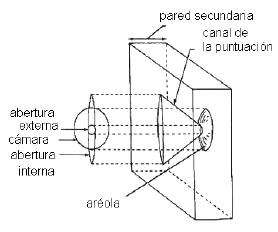
Son aquellas en las que la pared secundaria, al depositarse, hace un
reborde o aréola formando la cámara de la puntuación que se abre al lumen celular a través de la
abertura de la puntuación. La forma de la última puede concordar o no con el contorno de la aréola. Son de estructura más compleja y más variada que las simples. Se presentan principalmente en fibrotraqueidas y elementos conductores del xilema
(Fig. 7.21).
Cuando la pared secundaria es muy gruesa, se puede
diferenciar además de la cámara, el
canal de la puntuación, con la abertura interna hacia el lumen de la célula, y la abertura externa hacia la cámara de la puntuación.
El canal puede tener forma
de embudo aplanado, y entonces las aberturas interna y externa
difieren: la interna es lenticular o lineal y la externa es pequeña
y circular. En un par de puntuaciones, las aberturas internas están
frecuentemente dispuestas en cruz, en relación con la disposición
inclinada de las fibrillas de celulosa en la pared secundaria (Fig.
7.22).
Fig. 7.21,
Esquema
tridimensional de puntuación areolada
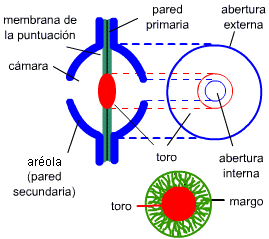
En las coníferas (gimnospermas) y algunas angiospermas (Oleaceae,
Ulmaceae, Thymeleaceae) las puntuaciones areoladas presentan toro, un engrosamiento central secundario lignificado.
La zona periférica llamada
margo está formada sólo por las fibrillas dispuestas
radialmente, ya que la matriz y la laminilla media han desaparecido por acción enzimática.
(Fig. 7.23). En las angiospermas la membrana de cierre no presenta aberturas visibles al microscopio, y ofrece así una resistencia mayor al transporte del agua.
En los elementos conductores, el paso del agua de una célula a otra se realiza a través de la puntuación interviniendo el toro en la regulación de dicho mecanismo.
Cuando el toro está en posición media, el agua pasa libremente a través del margo; cuando el toro se halla adosado a una de las aberturas de la puntuación, actúa como obturador impidiendo el paso del líquido
(Fig. 7.24).
Fig. 7.24, Puntuación areolada
funcional y aspirada
En algunas plantas hay
excrecencias muy elaboradas sobre la cara interna
de la aréola, y en este caso se habla de puntuación
vestida u ornamentada (Fig. 7.25).
La superficie interna está cubierta de verrugas
o protuberancias ramificadas. Son difíciles
de distinguir con microscopio óptico, pero
con MEB han sido encontradas en unas 30 familias,
entre las que figuran Leguminosas, Mirtáceas,
Crucíferas, Litráceas. El incremento
de superficie causado por las ornamentaciones mejora
la adhesión del agua a las paredes de los vasos,
y previene la cavitación (formación
de burbujas de aire o embolismos, catastróficos
porque interrumpen el flujo de la columna de agua).
Las puntuaciones vestidas son comunes en plantas que
viven en áreas secas.
Generalmente las puntuaciones en las paredes de células adyacentes se corresponden en posición, es decir que se trata de dos puntuaciones enfrentadas que constituyen un
par de puntuaciones. En este caso la membrana de cierre está constituida por la laminilla media compuesta. Si ambas puntuaciones son simples, tenemos un
par de puntuaciones simples; si son areoladas, tenemos un par de puntuaciones
areoladas. Cuando una puntuación simple se enfrenta con una areolada, tenemos un
par de puntuaciones semiareolado.
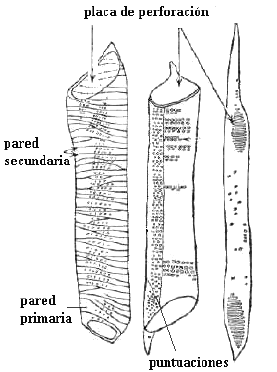
Es otro tipo de comunicación intercelular, en el que hay una interrupción de la pared primaria y laminilla media, además de la discontinuidad de pared secundaria.
Se presenta en células de los tejidos de conducción, en los vasos del xilema, donde constituyen las
placas de perforación (Fig. 7.26).