|
|
|
Tema 13: EPIDERMIS
13.5. Células
Oclusivas
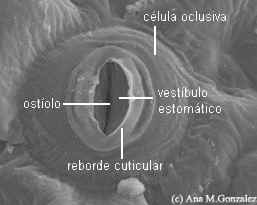
Las
de dicotiledóneas, Gimnospermae, muchas
monocotiledóneas y Pteridophyta son
arriñonadas, en forma de salchicha o banana en vista
superficial, con extremos redondeados. Suelen presentar
superficialmente un reborde cuticular externo que a veces
forma una verdadera cúpula o vestíbulo
estomático.
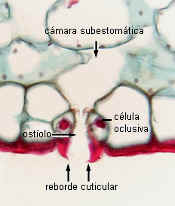
En corte transversal los
lados de este reborde se observan como
salientes
en forma de cuernos hacia la cara externa.
La cutícula se extiende sobre el poro y a veces también
forma salientes sobre la cara interna, constituyendo un vestíbulo
interno, como se observa en el corte de Clivia. Incluso
tapiza las células que limitan la cámara subestomática.
|
|
Pelargonium,
malvón:
estoma en vista superficial (MEB) |
Clivia:
estoma en corte (MO) |
 |
 |
|
Imagen de
Mauseth weblab |
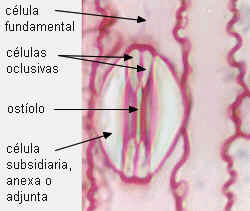
En Gramineae
y las células oclusivas son alargadas, con extremos
bulbosos donde el protoplasto de ambas células se halla
conectado por poros tan grandes que pueden ser atravesados por
mitocondrias o cloroplastos (Louguet, 1990). Estos poros se
originan por un desarrollo incompleto de la pared, es decir
que ambas células constituyen una unidad fisiológica.
El núcleo es alargado, con extremos ovoides y
filiforme en el medio. Las paredes de los extremos bulbosos
son delgadas, y las de la región media son gruesas.
|
Estomas de Saccharum
officinarum, caña de azúcar, con MO y MEB
|
 |
 |
|
Estomas de
gramíneas en diferentes planos de corte |
Estoma de maiz en
transcorte |
 |
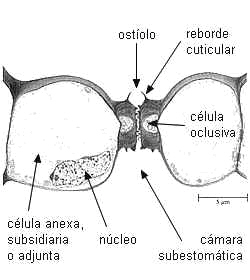 |
| Esquemas de
Strasburger 1994 |
Imagen de Raven
1999 |
Las Cyperaceae
tienen células oclusivas y núcleos con forma de pesas, igual
que las Gramineae,
pero la disposición de las microfibrillas de celulosa es
diferente y no hay datos disponibles sobre la presencia de
poros entre los extremos bulbosos (Sack
1994).
| En todos los casos, la pared que limita el
ostíolo es más gruesa que la pared opuesta, más delgada y
flexible que las paredes tangenciales.
Esta cualidad está relacionada con la capacidad de las
células oclusivas de cambiar de forma e incrementar su
volumen para controlar el tamaño de la abertura estomática.
La
disposición de las microfibrillas de
celulosa parecería estar en relación con esto: en
dicotiledóneas
la disposición es radial alrededor del poro; en las Gramineae es radial en los extremos bulbosos y en las células
anexas es paralela al ostíolo alargado.
|
Disposición de las
microfibrillas en la
pared celular de las células
oclusivas |
|
 |
|
Esquemas de Heath 1981 |
En muchas Gimnospermas
las paredes de las células oclusivas y de las células anexas
están parcialmente lignificadas, excepto en una banda que se
enfrenta al ostíolo. También
puede haber áreas lignificadas en estomas de algunas Criptógamas
y Angiospermae.
Las Flagellariaceae,
monocotiledóneas
emparentadas con las gramíneas, presentan estomas paracíticos
que al microscopio óptico son similares a los de gramíneas.
Las células oclusivas son apenas reniformes en vista
superficial, pero en corte longitudinal tienen forma de
caballete, con extremos bulbosos, de paredes engrosadas y sin
perforaciones entre ellos; los núcleos tienen sección elíptica.
Las células subsidiarias tienen una protuberancia que
se extiende por debajo de la porción angosta de las células
oclusivas, que forma una segunda abertura por dentro del ostíolo.
Tanto las células oclusivas como las subsidiarias
tienen paredes engrosadas diferencialmente, con varias capas.
Los orgánulos en ambas células parecen estar
confinados a determinados dominios.
Para otras familias como Palmae,
Rapateaceae, Marantaceae, Restionaceae se han descripto
estomas tipo gramíneas, faltan estudios detallados para
confirmarlo (Sack 1994).
| Las células oclusivas contienen
cloroplastos que acumulan periódicamente almidón, un núcleo
prominente y el sistema vacuolar más o menos dividido.
|
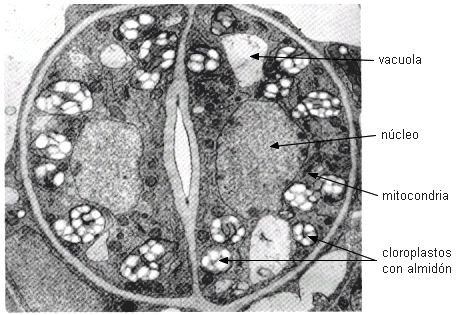 |
|
Imagen de Moore
1995 |
En
Selaginella las células
oclusivas son reniformes, y cada una recibe un solo plástido
de la célula madre que las origina. Inicialmente los microtúbulos
y los filamentos de actina se disponen radialmente con
respecto al ostíolo sobre las superficies periclinales
interna y externa. Después el citoesqueleto se reorganiza
sobre las paredes periclinales internas: los microtúbulos y
los filamentos de actina forman círculos concéntricos
alrededor del ostíolo mientras sobre las paredes externas se
mantiene la disposición radial (Cleary et al. 1993).
En
las células oclusivas los microfilamentos de actina cambian
su disposición en respuesta a estímulos fisiológicos y están
involucrados en los movimientos estomáticos. Fotografías obtenidas
con microscopio laser confocal y fluorescencia permitieron
comprobar que
cuando los estomas están abiertos
durante el día, los microfilamentos de actina están en el
citoplasma periférico, dispuestos radialmente con respecto al
ostíolo. Durante la noche, cuando los estomas están cerrados, los
microfilamentos están dispuestos al azar (Kim
et al. 1995).
Los movimientos estomáticos resultan de
cambios en la turgencia relativa de las células oclusivas y
anexas, causados por cambios en el potencial osmótico (Heath,
1981).
Se ha
constatado que los plasmodesmos entre las células
oclusivas y las vecinas están interrumpidos a la madurez del
estoma; la ausencia de comunicaciones intercelulares con las
células vecinas es lo que les permite funcionar
independientemente para controlar su turgencia (Willmer &
Sexton, 1979).
|
Commelina:
disposición de los microfilamentos de actina cuando
los estomas están abiertos |
Phaseolus vulgaris:
porción de célula oclusiva. Abajo izq.: detalle de
plasmodesmo obturador |
|
 |
|
|
Imagen
de Kim
et al. 1995 |
Imagen: Willmer y Sexton 1979 |

  
[ Tema 1 ] [ Tema 2 ] [ Tema 3 ] [ Tema 4 ] [ Tema 5 ] [ Tema 6 ] [ Tema 7 ] [ Tema 8 ] [ Tema 9 ] [ Tema 10 ] [ Tema 11 ] [ Tema 12 ] [ Tema 13 ] [ Tema 14 ] [ Tema 15 ] [ Tema 16 ] [ Tema 17 ] [ Tema 18 ] [ Tema 19 ] [ Tema 20 ] [ Tema 21 ] [ Tema 22 ] [ Tema 23 ] [ Actividades ] [ Tema 24 ]
|
|