|
|
Hoy se encuentran restringidas (bueno lo de restringidas, si se lee mas adelante, ya no parece un termino aplicable) a hábitats marginales como fuentes termales, depósitos profundos de petróleo caliente, fumarolas marinas, lagos salinosos (incluso en el mar Muerto...). Por habitar ambientes "extremos", se las conocen también con el nombre de extremófilas. Se considera que las condiciones de crecimiento semejan a las existentes en los primeros tiempos de la historia de la Tierra por ello a estos organismos se los denominó arqueobacterias (del griego arkhaios = antiguo). Membranas de las Archeae
Los lípidos presentes en las membranas son únicos desde el punto de vista químico, a diferencia de los eucariotas y las bacterias, en que los enlaces éster son los responsables de la unión entre los ác. grasos y glicerol, los lípidos de las Archaea poseen enlaces ÉTER para la unión del glicerol con cadenas laterales hidrofóbicas. En lugar de ac. grasos poseen cadenas laterales formadas por unidades repetitivas de una molécula hidrocarbonada como el isopreno. Los principales tipos de lípidos son los diéteres de glicerol. En algunos éteres las cadenas laterales (fentanil) se unen entre sí por enlaces covalentes formando una monocapa en lugar de la bicapa característica de las membranas, siendo más estables y resistentes, siendo habituales por lo tanto en las hipertemófilas. Paredes celulares
Algunas arqueobacterias metanogénicas poseen la pared celular formada por un compuesto similar al peptidoglicano de las bacterias, por lo que denomina pseudopeptidoglicano, con enlaces glucosídicos 1,3 en lugar de los 1,4 de los peptidoglicano. En otras archaeas la pared se compone de polisacaridos, glicoproteínas o proteínas. El tipo de pared más común es la capa superficial paracristalina (capa S) formada por proteína o glucoproteína, de simetría hexagonal. La pared celular impide la lisis celular y le confiere la forma a la célula. Las paredes de las Archaea son resistentes naturalmente a la lisozima, debido a la ausencia de peptidoglicano. La única arqueobacteria que carece de pared es Thermoplasma. Árbol Filogenético
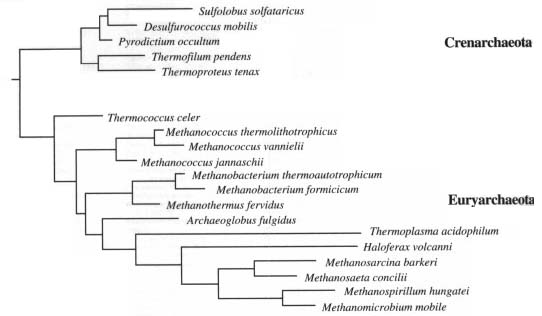
de Archaea
Sobre la base del análisis de la subunidad pequeña del ARN, las Archaea consisten en dos grupos filogenéticamente diferentes: Crenarchaeota y Euryarchaeota. Se diferencias por el tipo particular de ARN que presentan y por el ambiente en que habitan. Las Crenarchaeota (crenotas) es un grupo fisiológicamente homogéneo de hábitats enteramente termofílicos. en cambio las Euryarchaeota (euryotas) son un grupo fenotípicamente heterogéneo, que incluye a las metanogénicas, halófilas, etc. Basados en su fisiología se distinguen:
Además de las características unificadoras de las arqueobacterias, (pared celular sin mureína, lípidos de membranas con enlaces éter, etc.), estos procariotas exhiben atributos bioquímicos que le permiten adaptarse a estos ambientes extremos. Las Crenarchaeota son principalmente hipertermofílicos dependientes del sulfuro y los Euryarchaeota son metanogénicos y halófilos extremos. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Los procariotas Metanogénicos son habitantes normales del rumen de vacas y rumiantes. Una vaca puede eliminar unos 50 litros de gas metano por día, en el proceso de eructación. El metano es un importante gas del efecto invernadero que se acumula en la atmósfera a velocidad alarmante. Cuando se destruyen áreas verdes y se reemplazan por ganado se produce un doble impacto en el efecto invernadero ( "double-hit"):
|
Por otra parte gran cantidad de gas metano es producido durante el tratamiento de los líquidos cloacales, sin embargo normalmente es descartado en lugar de ser reciclado.
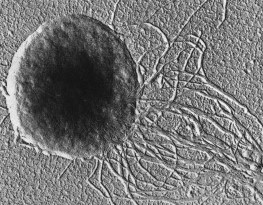
Methanococcus jannischii
Methanococcus jannischii fue originalmente aislada de una muestra tomada de una chimenea ( white smoker: fumarola blanca) a 2.600 metros de profundidad en el Pacífico Este. Puede crecer en un medio de cultivo mineral que contenga solo H2 y CO2 como fuente carbonada y en un rango de temperatura entre 50º - 86º grados. Estas células son cocos irregulares móviles, debido a la presencia de dos haces de flagelos polares insertos cerca del mismo polo .
Viven en ambientes naturales como el mar Muerto, el Great Salt Lake (Colorado USA), o en estanques de evaporación de agua salada, donde la concentración de sal es muy alta (hasta 5 molar o 25 por ciento de NaCl). Estos procariotas requieren la sal para el crecimiento, sus paredes celulares, ribosomas y enzimas se estabilizan con el ión Na+. Halobacterium halobium, la especie predominante en Great Salt Lake, se adapta al ambiente altamente salino por el desarrollo de una "membrana púrpura", que toma esta coloración por la presencia del pigmento del tipo de rodopsina llamado bacteriorodopsina que reacciona con la luz formando un gradiente de protones a lo largo de la membrana que permite la síntesis de ATP. Este es el único ejemplo en la naturaleza de una fotofosforilación sin clorofila. Estos organismos son heterótrofos y aerobios, la alta concentración de ClNa en el ambiente limita la disponibilidad de O2 para la respiración, por lo que usando bacteriorhodopsina aumentan su capacidad de producir a ATP, convirtiéndolo a partir de la energía lumínica.
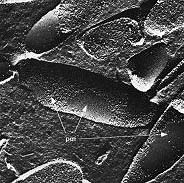
Halobacterium salinarium
Halobacterium salinarium es una halofila extrema que crece a 4 - 5 M NaCl y no crece por debajo de 3 M NaCl. La preparación por criofractura muestra la estructura de la superficie de la membrana celular y revela pequeños parches de "membrana púrpura" que contienen el pigmento bacteriorrodopsina embebidas en la membrana plasmática, este pigmento expulsa un protón de la célula, creando así un gradiente de protones que puede ser usado para generar ATP.
Representan varias líneas filogenéticas de Archaea. Estos organismos requieren temperaturas muy altas (80º - 105º grados) para crecer. Sus membranas y enzimas son inusualmente estables a estas temperaturas. La mayoría de ellas requiere sulfuro para crecer, algunas son anaerobias y usan el sulfuro como aceptor de electrones en la respiración, en reemplazo del oxígeno. Otras son litotróficas y oxidan sulfuro como fuente de energía, crecen a bajo pH (< pH 2) dado que acidifican su ambiente oxidando Su (sulfuro) a SO4 (ác. sulfúrico). Estos hipertermófilos son habitantes de ambientes calientes, ricos en sulfuro asociados a los volcanes, como los manantiales clientes, géiseres y las fumaroles del Parque National de Yellowstone , en respiraderos termales ("smokers") y en fracturas del piso oceánico. Sulfolobus fue el primer Archeae hipertermofílicos descubierto por Thomas D. Brock de la Universidad de Wisconsin en 1970. Su descubrimiento, junto al de Thermus aquaticus en el Parque Yellowstone, iniciaron el campo de la biología de los hipertermófilos. Thermus aquaticus, (fuente de la enzima taq polimerasa usada en la reacción en cadena de la polimerasa , PCR), crece a 70º grados. Sulfolobus crece en ambientes rico en sulfuro, manantiales calientes, 90º grados y pH 1. Thermoplasma, también descubierta por Brock, es un termófilo único, ya que es el representante exclusivo de una línea filogenética de Archaea. Thermoplasma recuerda a las bacterias micoplasmas ya que carece de pared celular. Thermoplasma crece a 55º grados y pH 2; solo han sido encontradas en pilas calientes de carbón, los cuales son productos de desecho humanos.
 |
![Fuentes termales Yellowstone [11 -> 53 Kb]](figbac/lagop.jpg)
|
|
Sulfolobus acidocaldarius (T.D.
Brock) |
Parque National de Yellowstone |
Sulfolobus es un termófilo extremo que se encuentra en manantiales ácidos productos de calentamiento por volcanes, y suelos con temperaturas entre 60º - 95º gradosC, y pH 1 a 5.
 |
 |
| Yellowstone National Park, USA, izq: Octopus Spring, der: Obsidian Pool. http://www.ucmp.berkeley.edu/archaea/archaea.html | |
A pesar que las Archaea son extremófilos por excelencia, también pueden encontrarse Bacterias, e inclusive algunos eucariotas en estos hábitat. Ninguna bacteria produce metano, pero existen algunas que creen en estos ambientes. Con respecto a la tolerancia ácida, una bacteria: Thiobacillus, puede crecer a pH 0. Un alga, Cyanidium, también puede crecer a pH 0. En ambiente supercálidos ( > de 100º C), los Archaea son exclusivos. Ninguna bacteria puede crecer en altas concentración de sales.
| Archaea | Bacteria | Eukarya | |
| Pared | pseudopeptidoglicano, o solo por proteínas | peptidoglicanos | plantas (celulosa), animales (ninguna), fungi (quitina) |
| Membrana | Lípidos: las cadenas hidrocarbonadas ramificadas están unidas al glicerol por enlaces éter | lípidos: las cadenas de ac. grasos están unidas al glicerol por enlaces ester | |
| Genoma | ADN único, circular, presencia de plásmidos | ADN fragmentado en cromosomas múltiples | |
![]()
![]()
|
Arqueobacterias (del griego arkhaios = antiguo; bakterion = bastón: grupo de procariotas de unos 3.500 millones de años de antigüedad, presentan una serie de características diferenciales que hicieron que Carl Woese, profesor de la Universidad de Illinois, Urbana, U.S.A., proponga su separación del reino Moneras y la creación de uno nuevo: Archaea, propuesta que hoy es cada vez mas aceptada. | |
|
ARN ribosómico: Uno de los tres tipos de ARN, el ARNr es un componente estructural de los ribosomas. Son el "core" (parte principal) de los ribosomas y posiblemente la clave del mecanismo de traducción de las proteínas. Su estudio comparativo llevó a postulación de un Árbol Filogenético Universal. | |
|
ATP(adenosín trifosfato): El principal producto químico utilizado por los sistemas vivientes para almacenar energía, consiste en un una base (adenina) unida a un azúcar (ribosa) y a tres fosfatos. |
![]()
![]()
| Brown, J.W. et al, Gene Structure, Organisation and Expression in Archaebacteria. CRC Critical Reviews in Microbiology, Vol 16, No. 4, 1989 | |
| Balows, A., H.G. Truper, M. Dworkin, W. Harder, and K.-H. Schleifer (eds.). The Prokaryotes, 2nd ed. Springer-Verlag, New York. 1992. Published in four volumes. The most complete reference on the characteristics of prokaryotes. Includes procedures for the selective isolation and identification of virtually all known prokaryotes. | |
| Danson et al., Archaebacteria, Biochemistry and Biotechnology, London: Biochemical Society, 1992 | |
| Holt, J.G. (editor-in-chief). Bergey's Manual of Systematic Bacteriology. Volume 1, 1982. Gram-negative bacteria of medical or industrial importance. Volume 2, 1986. Gram-positive bacteria of medical or industrial importance. Volume 3, 1988. Other Gram-negative bacteria, cyanobacteria, Archaea. Volume 4, 1988. Other Gram- positive bacteria. This is the standard authoritative guide to bacterial taxonomy and identification. This the usual place to begin a literature survey or an identification process of a specific bacterial group. | |
| Madigan, M.T., J.M. Martinko and J. Parker. Brock Biology of Microorganisms, 8th ed. Prentice-Hall, Inc., Englewood Cliffs, New Jersey. 1997. | |
| M.Ciaramella et al, Molecular biology of extremophiles, World Journal of Microbiology and Biotechnology, Vol 11, pp 71-84, 1995 | |
| Raven, P.H., R.F. Evert and S.E. Eichhorn. Biology of Plants, 5th ed. Worth Publishers, New York. 1992. | |
| Stanier, R.Y., J.L. Ingraham, M.L. Wheelis, and P.R Painter. The Microbial World, 5th ed. Prentice-Hall, Inc., Englewood Cliffs, New Jersey. 1986. |
Material traducido y modificado de Kenneth Todar University of Wisconsin-Madison Department of Bacteriology, por Dra. Ana Maria Gonzalez
martes, 05 de noviembre de 2013
|