|
|
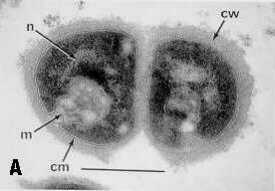
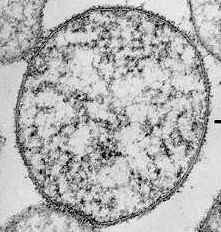
La relación topográfica entre la pared celular y la membrana plasmática se observa en la microfotografía electrónica de Micrococcus lysodeikticus (A) y de Bacteroides melaninogenicus (B).
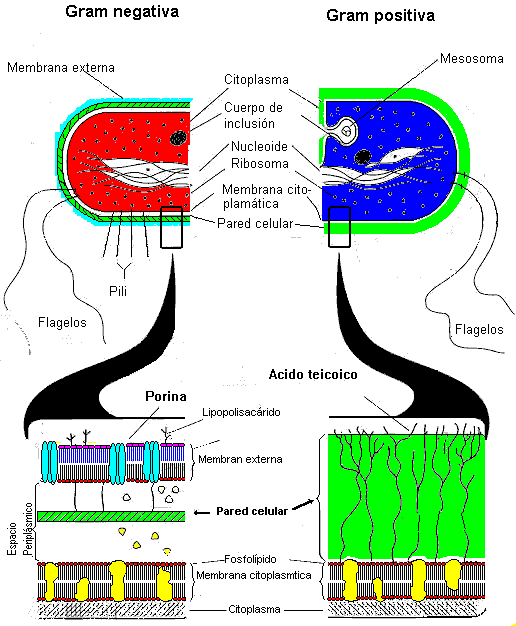
Tanto las bacterias Gram positivas como las Gram negativas poseen una pared celular de peptidoglicanos que les confieren su forma característica y les provee de protección mecánica. Dos grupos de bacterias carecen de pared celular:
La pared celular bacteriana rodea al protoplasto. La pared bacteriana es permeable a las sales y a muchas sustancias de bajo peso molecular. No es rígida sino elástica como el "cuero" de una pelota de fútbol, el protoplasto bacteriano confiere una cierta rigidez al conjunto protoplasto + pared que lo rodea, derivada de su presión interna o turgencia (al igual que la cámara hinchada de la pelota). La presión interna del protoplasto esta determinada osmóticamente. Se debe tener en cuenta que la membrana citoplasmática es la verdadera barrera osmótica (es semipermeable y controla la entrada y salida de sustancias a la célula). La pared celular y la plasmólisis
|
Sus principales características son:
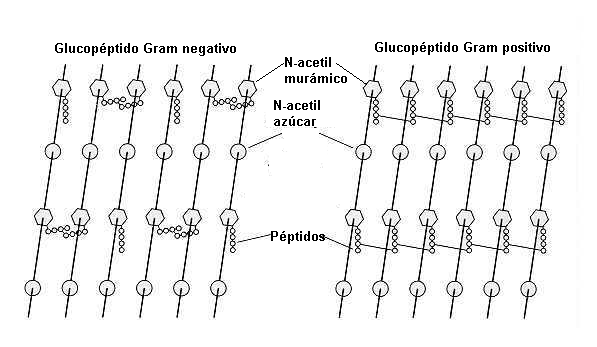
| La red de mureína esta muy desarrollada y llega a tener hasta 40 capas | |
| Los aminoácidos implicados varían de una especie a otra. | |
| La constitución del esqueleto es característica de la especie y constituye una buen parámetro taxonómico | |
| Es frecuente la presencia de los aminoácidos L-diaminopimélico o de lisina | |
| Los polisácaridos están unidos por enlaces covalentes (en el caso de tenerlos) Su contenido proteico es bajo. | |
| En ella se encuentran ácidos teicoicos |
| La red de mureína presenta una sola capa | |
| La constitución del saco de mureína es igual en todas las bacterias Gram negativas. | |
| Contiene siempre únicamente meso-diaminopimélico | |
| Nunca contiene lisina | |
| No se encuentran puentes interpeptídicos. | |
| Se encuentran grandes cantidades de lipoproteínas y lipopolisacáridos que representan hasta el 80 % del peso seco de la pared celular. Para mantener la estabilidad de las capas de lipopolisacáridos es necesario el ión Ca++. | |
| En las bacterias Gram negativas la capa de mureína puede ser atacada por la lisozima cuando se las trata con EDTA (Etilen-diamino-tetracético). Este agente, al quelar el Ca++ libera una parte de los lipopolisacáridos y permite la acción de la enzima.. | |
| Hasta ahora no han podido demostrarse ácidos teicoicos. |
Un viejo refrán dice que Dios llama a la puerta de un hombre solo una vez en la vida. No es el caso de Sir Alexander Fleming a la de él llamo dos veces (o más), aunque es necesario coincidir que no es suficiente con el llamado, amén de él, es necesario escuchar y responder.
Según se cuenta el descubrimiento de la lisozima tuvo lugar cuando las lagrimas de un niño (que estaba en el laboratorio donde trabajaba Fleming), caen en un tubo y aclaran una suspensión Micrococcus lysodeikticus, que, hoy se sabe, es la bacteria mas sensible a la lisozima, ma siii....
Este hecho esta un poco menos difundido que la novelesca historia de la espora de Penicilium que entro por la ventana del laboratorio de Fleming, cayo en una placa de Petri y al crecer puso de manifiesto la acción bactericida del antibiótico que llamo penicilina....( Florey y Chain en Oxford la aislaron, dieron las pautas de su producción industrial y al hacerlo pusieron en marcha la era de los antibióticos, por ello, Fleming , Florey y Chain recibieron el premio Nobel en 1945 )
La lisozima descubierta en 1922, es una enzima que rompe el enlace beta glucosídico de la mureína. Se la encuentra en el líquido lagrimal, secreciones nasales y en la clara de huevo. También se la ha aislado de bacterias y bacteriófagos. La acción de la lisozima se pone en evidencia por un aclaramiento rápido de una suspensión bacteriana, Micrococcus lysodeikticus ya se lisa con 1 ug de lisozima / ml. Para lisar otras bacterias p.e Bacillus megaterium se necesitan 50 ug / ml . La capa de mureína de muchas bacterias Gram negativas solo es atacada por la lisozima cuando se añade EDTA (Etilen-diamino-tetracético).
El mecanismo de lisis es el siguiente: la destrucción de la pared celular deja al protoplasma de las bacterias rodeado únicamente por la membrana celular ("protoplasto"), lo cual convierte a la bacteria en un organismo extraordinariamente sensible a las variaciones de tonicidad del medio, esta es la base del fenómeno de aclaración que tiene lugar luego de la acción de la lisozima, cuando la solución es hipotónica.

Los protoplastos son estables en medios hipertónicos e isotónicos; en medios hipotónicos, tal lo señalado, estallan y dejan restos de membrana citoplasmática llamados "fantasmas" (ghosts).
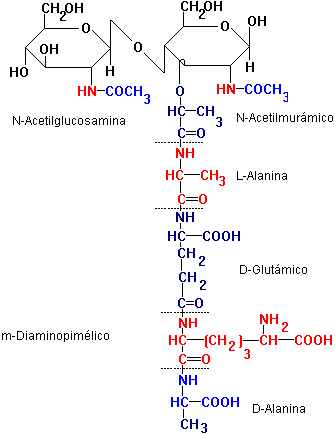
El proceso de síntesis de la pared celular comienza en el citoplasma bacteriano, a partir de N-acetilglucosamina-1-fosfato que se une al UDP (uridin difosfato).
| El UDP se combina con la N-acetilglucosamina-1-fosfato, para dar UDP-N-acetilglucosamina que primero forma el éter láctico y luego en sucesivos pasos enzimáticos se unen al mismo los cinco aminoácidos para formar el N-acetilmurámico. |
|
|
| En el siguiente paso, que ocurre en el ámbito de la membrana citoplasmática la molécula que es hidrófila cambia a lipófila, lo cual facilita su transporte, por el cambio del UDP por undecaprenil-fosfato y el agregado de un pentapéptido de glicina a nivel de la L-lisina | |
| En la siguiente fase, en el ámbito de la pared celular, se produce la transglucosidación y formación del enlace beta alargando de esta manera la molécula. | |
| Los enlaces transversales entre moléculas del polímero se produce por transpeptidación, se libera una D-alanina y el grupo carboxilo se une a un grupo amino de la lisina de otro oligopéptido, también se libera el undecaprenil-fosfato. |
La penicilina interfiere en este ultimo paso, es decir impide la unión transversal o puente interpeptídico pero no la elongación del polímero. De la misma manera actúan las cefalosporinas, vancomicina, bacitracina y cicloserina. Debe notarse que la síntesis de la pared no se realiza cuando no hay lisina disponible o cuando se impide la racemización de la L-alanina y por lo tanto la disponibilidad de la D-alanina por efecto de la c-clicoserina (oxamicina).

En la bacterias Gram negativas se observa por fuera del saco de mureína una capa semejante a la membrana celular por lo que se ha dado en llamarla membrana externa, es de composición compleja y en ella intervienen fosfolípidos, proteínas y lipopolisacáridos.
Cumple funciones mecánicas y fisiológicas. En esta bicapa lipídica (compuesta por el lípido A del lipopolisacárido en su parte externa y los fosfolípidos en la interna) se encuentran proteínas que la atraviesan en todo su espesor y que delimitan poros (por eso se denominan porinas) hidrófilos que permiten el paso de sustancias hidrófilas de bajo peso molecular. No se encontraron sistemas de transporte activo.
La membrana externa se encuentra unida al peptidoglicano a través de la proteína conocida como lipoproteína de Braun cuya parte lipídica se encuentra en la membrana y la proteína unida covalentemente a la mureína. de la membrana. La membrana externa tiene unos puntos de contacto con la membrana interna que se denominan uniones de Bayer y que se supone son importantes en el transporte a través de la membrana.
ESPACIO PERIPLÁSMICO
En las bacterias Gram negativas el espacio que va desde membrana plasmática a membrana externa (ver figura abajo) se denomina espacio periplásmico, tiene aparentemente una consistencia gelatinosa.
| En numerosas bacterias contiene abundantes enzimas, por ejemplo aquellas que inician la degradación de substratos como la glucosa, o la transformación de compuestos inorgánicos como los nitratos. | |
| También se encuentran las depolimerasas que actúan sobre los biopolímeros (proteasas, polisacaridasas, nucleasas y otros). | |
|
Otras enzimas importantes presentes en el espacio periplásmico son las b-lactamasas responsables de la destrucción de los antibióticos b-lactámicos y, por tanto, de la resistencia de ciertas bacterias a ellos. | |
|
Asimismo se encuentran allí sensores químicos destinados a detectar variaciones ambientales. |
Algunas de estas enzimas están libres y otras ligadas a la membrana citoplasmática.
En las bacterias Gram-positivas no hay, en realidad, espacio periplásmico; pero pareciera que la parte más interna del peptidoglicano puede desarrollar una función similar reteniendo mediante fuerzas electrostáticas moléculas de enzimas equivalentes a las periplásmicas de las Gram-negativas.
Los lipopolisacáridos
Los lipopolisacáridos con una estructura característica se encuentran en la capa externa, a diferencia de la membrana plasmática los forman:
La parte glucosídica corresponde a dos molécula de N-acetilglucosamina (es, para ponerlo en un nombre mas largo una glucosaminadisacárido, es la estructura equivalente al
|
glicerol de los fosfolípidos) sus grupos hidroxilos están esterificados por ácidos grasos C12, C16, C18, que al ser hidrofóbicos se orientan hacia el interior. Se lo conoce como lípido A
| unida a ellas y proyectándose al exterior se encuentra una cadena glicosídica. Esta cadena posee una zona central R y a ella le sigue extermamente la cadenas heteropolisacárida O- especifica que representa a los antígenos somáticos. |
La función del lipopolisacárido en las bacterias es la de incapacitar las defensas del huésped, proporcionar carga negativa a la superficie de la membrana y estabilizarla.
Constituyen una de las endotoxinas más activas de las bacterias, son fuente de origen de fiebre, diarrea y provocan una fuerte respuesta inmune, siendo uno de los agentes responsables del llamado «shock endotóxi producido cuando las células del sistema inmune entran en contacto con él. Tienen gran importancia en el diagnóstico bacteriológico y en la identificación de infecciones.
| Distintas cepas se diferencian entre sí por las llamadas cadenas laterales O específicas ya señaladas, hecho que puede ponerse de manifiesto por métodos inmunológicos. | |
| Las células bacterianas crecen sobre agar generalmente como colonias lisas y brillantes denominadas formas S (por smooth= lisos); la superficie regular contiene mucha agua debido a la presencia de las cadenas polisacáridas O-específicas. | |
| Estas formas lisas mutan espontáneamente a formas que crecen como colonias planas y rugosas, denominadas formas R (por rough = rugoso). | |
| Cuando invaden un organismo, la presencia de esas cadenas de polisácaridos dan a las bacterias S una ventaja selectiva al ser más resistentes a la fagocitosis por los leucocitos y por ello más virulentas. | |
| Hasta que el hospedador no forme anticuerpos y éstos se unan a los polisácaridos, las bacterias no son atacables, la gran variedad de polisacáridos O-específicos en bacterias patógenas puede deberse a una selección de tipos O-antigénicos (mutantes) nuevos cada vez; tienen por tanto una ventaja en el desarrollo, porque el hospedador no puede disponer simultáneamente de los anticuerpos contra cientos de antígenos. |
![]()
![]()
| Acidos teicoico: son cadenas de moléculas de glicerina o ribitol esterificadas entre sí por puentes fosfato, portan una fuerte carga negativa (probablemente sirvan para "secuestrar" cationes). Los ácidos teicoicos se unen probablemente a la mureína a través del fosfato formando una amida. Son muy antigénicos. Los grupos sustitutivos de los polioles incluyen D-alanina (como éster), N-acetilglucosamina, N-acetilgalactosamina, los sustituyentes son característicos de las especies bacterianas. 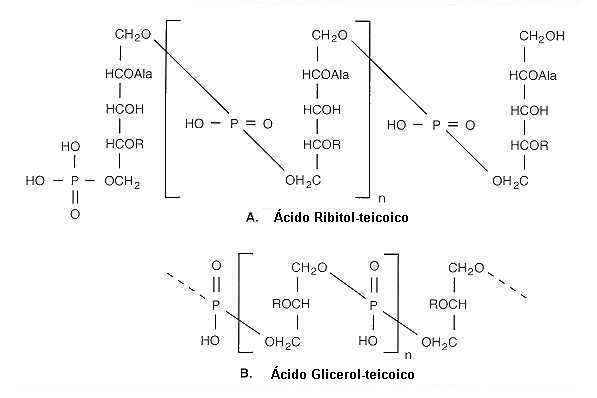 |
Estructura de los ácidos teicoicos de la pared celular
(A) Acido Ribitol- teicoico con unidades repetitivas unidas por enlaces 1,5-fosfo diéster de D-ribitol y éster de D-alanina en posición 2 y sustituyentes glicosidícos en posición 4 (R= N-acetilglucosamina en S. aureus o alfa-glucosil en B. subtilis .
(B)Acido Glicerol- teicoico con unidades repetitivas unidas por enlaces 1,3-fosfo diéster (1,2,en algunas especies). (R =H o D-alanina o
glucosil).
| Pelota de fútbol: En mi época la "numero 5" tenia un "cuero con tientos" y una cámara. Con un inflador se inflaba la pelota hasta que adquiría la consistencia debida y, salvo que ahora las pelotas no tienen tientos y otras sofisticaciones, para que tengan esa consistencia hay que inflarlas. | |
| El valor osmótico del contenido celular es equivalente al de una disolución al 10-20% de sacarosa; . Normalmente la concentración de azúcares y sales es superior en el interior de las células que en el exterior y la célula capta tanta agua como lo permite la pared celular | |
| Aquíles: héroe de la mitología griega. La leyenda cuenta que su madre lo sumergió en la laguna Estige lo que lo hizo invulnerable, excepto en el talón por donde lo sostuvo. Muere a consecuencia de una flecha que París, el héroe troyano, raptor de la bella Helena, acertó en su talón. Por extensión "talón de Aquiles" = punto débil. | |
| Tinción de Gram: |
- El procedimiento se inicia fijando a la llama las bacterias extendidas en un portaobjetos.
- Las mismas se tiñen con una solución del colorante básico cristal violeta.
- El paso siguiente consiste en un tratamiento con solución de Lugol (iodo/ioduro de potasio). El yodo forma una laca con el cristal violeta, que es insoluble en agua y soluble en alcohol o acetona
- El preparado se trata después con alcohol o acetona
- Finalmente se procede a realizar una coloración de contraste (diferenciación) con otro colorante (fucsina).
- Como consecuencia de este tratamiento las bacterias que retienen el complejo colorante-yodo durante el paso 4 quedan azules y se denominan Gram positivas.
- Las células que NO retienen el complejo colorante-yodo durante el paso 4 al tomar el colorante de contraste (paso5) quedan rojas y se denominan Gram negativas.
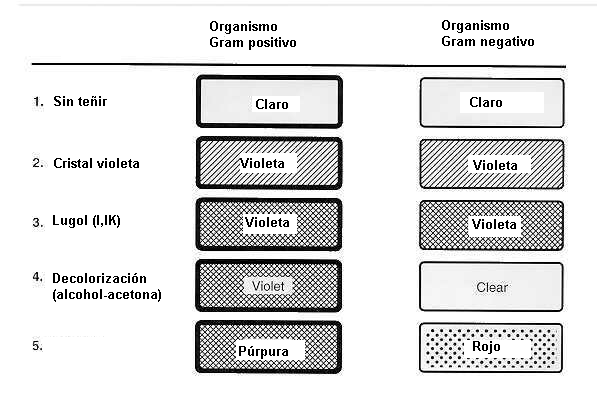 |
| Secuencia de las etapas de la coloración de Gram y la coloración resultante en las Gram negativas y positivas. |
Redacción y diagramación a cargo de: Dr. Jorge S. Raisman. Revisado en Setiembre de 2005.
|