|
|||||||||
|
|
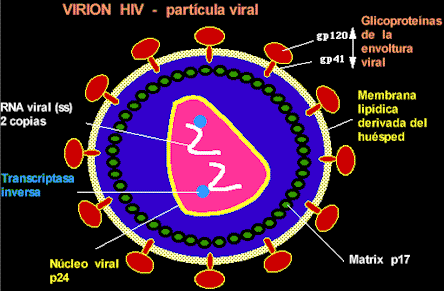
Virión VIH-1 Estructura del virión: La parte central de la nucleocápside contiene dos copias del genoma viral (ARNss), proteínas del núcleo y transcriptasa inversa. La envoltura que lo rodea está compuesta de lípidos (derivados de la célula huésped), y glicoproteínas virales (gp120 y gp41).
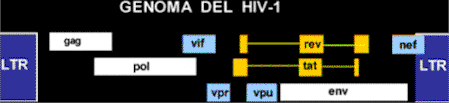
Genoma del VIH-1 Genes estructurales Genes de transactivación: genes esenciales
para la replicación in vitro: Genes accesorios: genes no esenciales para
la replicación viral in vitro. Características principales del genoma VIH-1
Proteínas del ViriónProteínas de la cápside (núcleo del virión)Las proteínas de la cápside son productos del gen gag. Los productos gag incluyen: la Matrix (MA), la Cápside (CA), y las proteínas de la nucleocápside (NC), que derivan de la proteolísis de la poliproteína Gag, Pr55gag.. La fragmentación está mediada por la proteasa viral (PR), la cual es el objetivo de numerosos fármacos antivirales. MA (proteína matrix): Se cree que el MA interacciona con las proteínas transmembranosas de la envoltura viral (TM). Una interacción que puede ayudar a reclutar las proteínas necesarias para construir el núcleo viral durante el ensamblaje viral. CA (proteína de la cápside): Es muy importante para el ensamblaje de la nucleocápside. CA (y el precursor Gag Pr55gag) también interacciona con las proteínas celulares, como la ciclofilina A (CyA = cyclophilin A) que es incorporada en los viriones VIH-1 y se necesita para la capacidad infecciosa del virus. Se piensa que la CyA facilita la decapación de la cápside del VIH-1 en las etapas tempranas de la infección, probablemente al desestabilizar la proteína CA del complejo viral nucleoproteico (CyA es un chaperón molecular que puede alterar el moldeamiento de las proteínas). NC (proteína de la nucleocápside): Es una proteína básica que se une (de modo no específico) al ARN viral. Puede contribuír a condensar el ARN viral para que pueda ser almacenado dentro de la nucleocápside. NC también se une de modo específico, a la mayor señal de enpaquetamiento del genoma del VIH-1: la señal psi ó y.
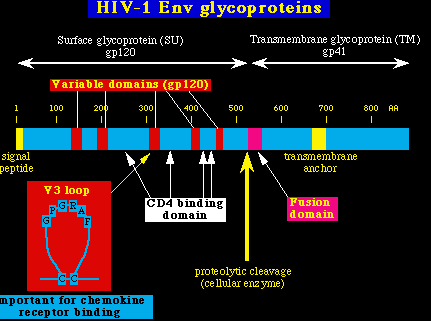
Los enzimas virales son productos del gen pol. Pol codifica las enzimas: proteasa (PR), transcriptasa inversa (RT), la ARNsa H, y la integrasa (IN). Al igual que pasa con los productos del gen gag, éstas enzimas son hechas a partir de la fragmentación de grandes poliproteínas. En este caso, la proteína precursora es una poliproteína Gag-Pol, Pr160gag-pol, hecha a su vez como el producto de un cambio en la traducción del mARN viral, y tiene su sitio específico en la secuencia del ARN. (El Gag y el Pol se codifican en diferentes secuencias de lectura del genoma VIH-1, y las dos secuencias de lectura abierta se solapan. Anotar que la proteína Pr160gag-pol tiene un potencial autolítico. PR (proteasa). Es una enzima dimérica que divide específicamente a la poliproteína Gag-Pol del VIH-1. Se ha definido tanto su estructura cristalina, como su especifidad de lugar; lo que permite el diseño de una gran variedad de inhibidores enzimáticos que puedan interferir con su unión al substrato, la dimerización del enzima ó la actividad enzimática. RT (transcriptasa inversa) y la ARNsa H. La transcriptasa inversa es una polimerasa DNA, dependiente del ARN, que sintetiza DNA a partir de una plantilla de ARN. También posee actividad ARNsa H, lo que la permite degradar el ARN de los híbridos ARN:DNA (Es un paso necesario durante la transcripción inversa). Ha sido estudiada la estructura cristalina de la RT y se han logrado producir varios inhibidores específicos de ésta enzima. Una de las propiedades más importantes de la RT es que no tiene actividad de corrección de lectura. Por ende, no puede corregir los errores que hace durante la síntesis de DNA, y así la replicación del VIH-1 tiene una frecuencia alta de mutaciones (lo mismo que le pasa a otros retrovirus). IN (Integrasa). La IN cataliza la inserción de la doble cadena linear del DNA del VIH-1 en los cromosomas celulares. Es un paso obligado en el ciclo vital del VIH-1, y por consiguiente la IN es una enzima que puede servir como blanco potencial de futuros fármacos antivirales. Glicoproteínas de la envoltura viral Las glicoproteínas de la envoltura del VIH-1 son el resultado de la acción del gen Env. Entre éstas glicoproteínas están: la glicoproteína de la superficie de la envoltura (SU ó gp120) y la glicoproteína transmembranosa (TM ó gp41). Lo mismo que ocurría con Gag y Pol, éstas glicoproteínas se producen por fragmentación de grandes glicoproteínas precursoras (gp160). La fragmentación está catalizada por una proteasa celular.
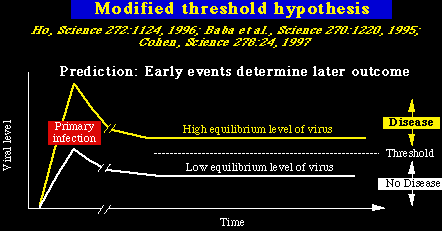
SU ó gp120 (glicoproteína de superficie). Es una proteína muy glicosilada que se localiza fuera de la partícula viral. No tiene acción transmembranosa, y se une de modo no-covalente (se une débilmente) a la glicoproteína TM/gp41. Las dos glicoproteínas: gp120 y gp41 se encuentran como oligómeros dentro de la partícula viral (así: gp120 interacciona tanto con la gp41 como con otras moléculas gp120, y la gp41 hace otro tanto). En general, las secuencias codificadas por la gp120 de las diferentes cepas de VIH-1, son muy variables, y éstas variaciones de las secuencias pueden ser atribuídas a 5 regiones de cambio dispersas con las regiones conservadas. La tercera región variable contiene un segmento importante conocido como el asa V3. Esta asa de aproximadamente 30 aminoácidos está formada por dos resíduos de cisteína (unidos por un puente disulfuro), y tiene aminoácidos intactos (GPGRAF) en su cresta. El resto del asa es más variable, se ha demostrado que las variaciones del asa V3 altera el alineamiento del VIH-1 en la célula huésped, debido a su efecto en la interacción entre el gp120 y los receptores de las quinasas celulares. Lo más importante, el gp120 se une con una afinidad muy alta a la proteína celular CD4. Después de esta unión al CD4, el gp120 se une a los receptores de las quinasas celulares (notablemente el CCR5). Las regiones de unión del CD4 y los receptores de las quinasas del gp120, se conservan bastante bien, y se componen de múltiples regiones discontínuas que entran en contacto y se agrupan, debido a los pliegues de la proteína. TM ó gp41. (glicoproteína transmembranosa). El gp41 contiene residuos muy hidrofóbicos en un segmento corto de la parte N terminal (es el dominio de fusión). Esta región de fusión es importante porque permite que la membrana del virión se funda con la membrana celular y permita la entrada del virus en la célula. Además de la región de fusión, hay un segundo dominio hidrofóbico en la mitad del gp41, el cual sirve para anclar la proteínas a la doble capa de lípidos de la envoltura viral. Genes virales accesorios Genes no esenciales in vitro.Vif. Virion Infectivity Factor = Factor de Infectividad Viral. Es un factor importante en la maduración del virión, parece ser necesario para neutralizar un factor celular que inhibe el VIH-1 Vpu. Viral Protein, Unknown. = Proteína Viral Desconocida. Está presente en el VIH-1 pero ausente en el VIH-2. Disminuye la expresión del CD4 y promueve la gemación viral. Vpu también regula a la baja el complejo Clase I del MHC. Finalmente, Vpu tiene similitudes funcionales y estructurales con la proteína M2 del virus de la gripe A (que actúa con un canal iónico). Vpr. Viral Protein, Regulatory= Proteína Viral Reguladora. Detiene la proliferación celular en la fase G2 del ciclo celular, induce la diferenciación celular e interacciona con las proteínas celulares involucradas en la reparación del DNA. Ona consecuencia de la parada del ciclo celular en G2 inducida por el Vpr, es un aumento de la replicación viral, ya que el LTR viral es muy activo en dicha fase. Vpr también tiene una señal de localización nuclear (NLS) la cual es importante para la pre-integración nuclear del complejo VIH. En SIV, el Vpx tiene la función de la importación al núcleo que hace el Vpr, y es necesario para infectar los macrófagos. Vpx también es necesario para la diseminación viral depués de la primoinfección in vivo, sugiriendo que los macrófagos pueden ser muy importantes para una replicación precoz y la consiguiente propagación del virus (Hirsch et al. Nature Med. 4:1401, 1998). Nef. Negative factor (un nombre incorrecto).Su función fué un misterio por muchos años, pues se podía borar de los clones VIH-1 con escasos cambios en la actividad viral, en los estudios in vitro. Subsecuentemente, Nef fué localizado en las membranas celulares (por la adición de un lípido de cadena larga a la proteína, en un proceso conocido como miristoylación).Más tarde, se encontró que Nef, tenía unas propiedades potencialmente muy interesantes, como la de regular a la baja las moléculas CD4 y MHC clase I y la habilidad de activar las quinasas celulares, interfiriendo de ésta manera con los procesos celulares de señalización. Específicamente, Nef contiene el complejo peptídico PxxP, el cual permite que Nef se una al src, en la región de homologación 3 , dominio (SH3) de las quinasas de la tirosina celular, que forman parte de la familia src. La propiedad más dramática del Nef fué revelada gracias a un experimento magistral efectuado por Ronald Desrosiers and colleagues in 1991 (Kestler et al. Cell 65:651, 1991). Brevemente, borrar el gen Nef de un clon patogénico molecular del SIV convirtió al virus en inofensivo para los macacos adultos, estableciendo de este modo que Nef es indispensable para que el SIV pueda inducir la enfermedad in vivo. Los estudios hechos en el Nef del SIV han sido apoyados por los análisis de la Cohorte del Banco de Sangre de Sidney. Reveló que un grupo de 6 individuos que recibieron productos sanguíneos infectados con el VIH-1 de un único donante VIH-1 positivo, permanecieron en buen estado de salud por más de 10 años, sin caídas en los niveles de CD4. Análisis posteriores mostraron que la cepa de VIH-1 que infectó a éstas personas tenía una gran parte del gen Nef borrada Deacon et al. Science 270:988, 1995). Estos datos, junto con los resultados obtenidos por Desrosiers, sugieren que las cepas de VIH-1 con el nef borrado, pueden ser cepas atenuadas. El grupo de Desrosiers ha demostrado que la infección de macacos con cepas VIH-1, donde se ha borrado el nef, pueden proteger a ésto animales en contra de infecciones por cepas salvajes de SIV; lo que lleva a pensar que puede ser posible diseñar vacunas con virus vivos y atenuados contra el VIH. Cepas mejoradas, donde se han borrado múltiples genes, pueden permitir una vacuna con un balance óptimo de inmunogenicidad y de seguridad, y se planea un ensayo en humanos de una vacuna con cepas donde se han borrado los genes Nef, Vpr y Vpu.(http://www.iapac.org/). De todos modos, está claro que el VIH puede causar enfermedad en ausencia del nef. Unos pocos animales inoculados con cepas SIVDNef ó con cepas con triple deleción (SIVD3) , han desarrollado la inmunodeficiencia, y algunos monos recién nacidos inyectados con dosis altas de SIVDNef tmabién desarrollaron el SIDA. En términos generales, la posibilidad de causar la enfermedad ha sido relacionada con la habilidad del virus de replicarse in vivo a niveles elevados. esto ha llevado a la hipótesis del umbral, que en esencia estipula que el nivel precoz de replicación determina si la infección viral será patológica en un individuo cualquiera.
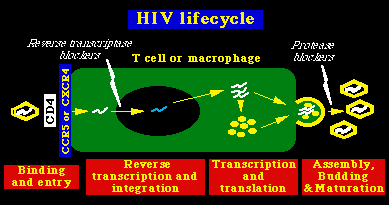
Etapas en la replicación del VIH-1
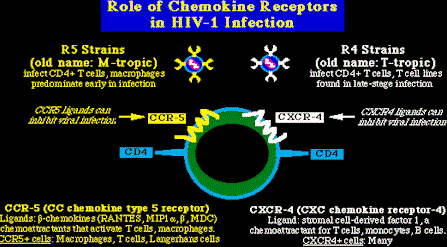
Unión del virión a la célula y su entradaEl paso inicial de la infección por el VIH-1, es la unión del gp120 a la molécula CD4 de la célula, paso obligado en la infección de las células T y de los macrófagos. esta interacción produce cambios de comportamiento tanto en el gp120 como en el gp41. En el gp120, el asa V3 es expuesta y de este modo el gp120 puede unirse a los receptores de las quimocinas, las cuales actúan como receptores auxiliares en la entrada del virión a la célula. La interacción del gp120 con la CD4 y los receptores de las quimocinas producen a su vez cambios en el comportamiento del gp41, el cual expone una zona de dominio de fusión hidrofóbica en la parte N-términus de la proteína, permitiendo la fusión de las membranas celular y viral. Hay que resaltar que los cambios de comportamiento pueden ser inhibidos por el péptido antiviral, T-20 (Nature Med. 4:1302, 1998). Cofactores de entrada: Aunque el CD4 es necesario para la entrada del virión, no es por si misma capaz de hacer que una célula sea infectada por el VIH-1 (por ejemplo, las células del glioma humano pueder ser manipuladas para que expresen CD4, pero no pueden ser infectadas por el VIH-1). Esto demuestra que el VIH-1 necesita un cofactor adicional para su entrada. Ed Berger y su equipo clonaron el primero de éstos cofactores en 1966, y el trabajo posterior de varios laboratorios lograron la identificación de dos grandes cofactores para la entrada del VIH-1, los dos son receptores de las quimocinas celulares.
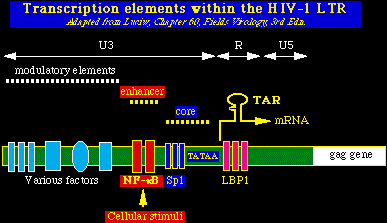
Transcripción inversa e integraciónDespués de la entrada del virión VIH-1 en la célula, las partículas del núcleo viral son liberadas y el ARN viral es convertido en una doble cadena linear de DNA por la transcriptasa inversa viral. El complejo DNA-núcleo viral (ó complejo de pre-integración) es transportado al interior del núcleo de la célula. Debido a que dicho complejo es muy grande para pasar de modo simple por los poros de la membrana nuclear, el proceso de importación al núcleo celular representa un estadío crítico en el ciclo vital del virus. El VIH-1 y otros lentovirus se diferencian de los oncoretrovirus, en que la importación al núcleo celular de los lentivirus es un proceso activo, que depende de la presencia de señales de localización nuclear dentro de algunas proteínas del virión. (en contraste, los complejos de pre-integración de los oncoretrovirus entran al núcleo solamente durante la mitosis, una vez que la membrana nuclear se ha disuelto). El resultado es que los lentivirus pueden infectar las células que no se dividen (como los macrófagos) y los oncoretrovirus no. Después de la importación al núcleo del complejo de pre-integración, el DNA linear del VIH-1 se integra en los cromosomas del huésped por medio de la integrasa, una enzima viral. Transcripción y traslaciónEl control de la expresión genética del VIH-1 se regula por medio de factores celulares y virales. El ARN viral se sintetiza en el núcleo, a partir del provirus VIH-1 que se ha integrado y su expresión está regulada por los elementos de control presentes en la región U3 de la larga secuencia terminal de repetición (LTR: Long Terminal Repeat). Los elementos de control incluyen:
Elementos de amplificación Incluyen sitios de unión para los factores de transcripción celular NFAT y NF-kB, los cuales actúan aumentando la expresión genética del VIH-1 en respuesta a la estimulación celular por diferentes mecanismos, como citocinas exógenas y activación de las células T. La facilidad del VIH-1 de responder a la activación celular es muy importante, pues la gran mayoría de las células T infectadas por virus están normalmente en reposo. Las condiciones que inducen la activación inmune aumentan la replicación viral. Este fenómeno puede explicar la aceleración del SIDA cuando hay otras infecciones presentes y los aumentos transitorios de la replicación del VIH-1 en las personas VIH-1+ vacunadas de modo rutinario.
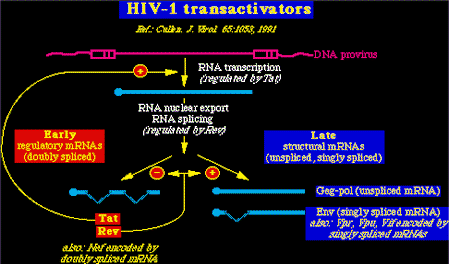
Trans-activadores viralesLos lentivirus en contraste con los onco-retrovirus, poseen transactivadores muy especiales, que pueden agruparse en dos categorías:
La acción combinada de estas proteínas reguladoras divide el ciclo vital de los lentivirus en dos fases temporales:
Hay que recordar que HTLV-I y II codifican proteínas análogas: Tax y Rex. Propiedades de Tat y Rev del VIH-1
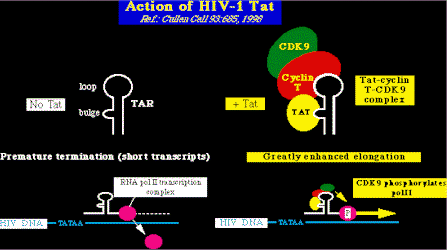
Acción del TatTat (trans-activator of transcription = trans-activador de la transcripción), es esencial para la replicación viral es una proteína corta (aproximadamente 86 aminoácidos). Tac actúa por medio de una estructura que está en una de las ramas del asa del ARN, en la región R del segmento LTR (muy cerca del sitio donde empieza la transcripción viral). A esta estructura se la conoce como TAR (the Tat-responsive element = elemento con afinidad al Tat), y Tat se une a una pequeña protuberancia dentro de esa estructura (proteínas celulares también se unen a este sitio). En general, Tat está formada por dos grandes regiones de control:
El principal efecto de la unión del Tat al Tar es aumentar la eficiencia de la elongación de la transcripción (por ejemplo: Tat interacciona con el complejo de la polimerasa II y aumenta su eficiencia). Este efecto es importante porque casi todas las secuencias transcritas del mARN del VIH-1 terminan de modo prematuro en la ausencia de Tat. Mirado de esta manera, se puede considerar al Tat como un complejo que evita la terminación de la transcripción (como la proteína N del bacteriófago lambda). Como aumenta Tat la eficiencia de la transcripción ? Tat actúa estimulando las quinasas celulares, que a su vez estimulan la eficiencia de la polimerasa II del ARN en los pasos iniciales de la elongación. Para hacer esto, Tat inteacciona directamente con la secuencia TAR del ARN y también con la ciclina T. La ciclina T a su vez, se une a una quinasa dependiente de la ciclina (CDK), CDK9, la cual es capaz de fosforilar la región C-terminal de la polimerasa II del ARN. El resultado es la hiper-fosforilación de los residuos de serina de la polimerasa II del ARN, con el efecto final de un auemnto de la eficiencia de la enzima.
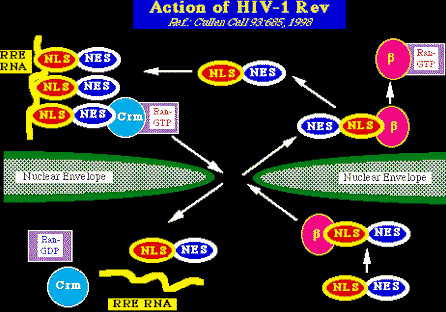
Acción del RevRev (regulator of viral expression = regulador de la expresión viral) es al igual que el tat, una proteína pequeña de aproximadamente 116 aminoácidos, esencial para la replicación viral. Como el Tat tiene regiones de dominiio bien definidas, entre ellas:
Secuencia final del Rev. Rev actúa por medio de un estructura compleja de ARN llamada RRE ( Rev-responsive element = elemento que reacciona con REV), el cual se encuentra en el gen env del VIH-1. La unión del Rev al RRE es necesaria para la exportación nuclear y la expresión del mARNs tardío del VIH-1 (por ejemplo: las secuencias del mARN sin unir y las que solo están unidas por un terminal, que codifican proteínas estructurales virales). Como con Tat y TAR, factores celulares también se unen al Rev y al RRE. Cómo funciona Rev ?: (see Cullen, B. Cell 93:685, 1998)
Tat y Rev: temas comunes
Actualizado el 1/99, Steve Dewhurst, Associate Professor of Microbiology
and Immunology URL: http://www.urmc.rochester.edu/smd/mbi/grad2/VIH99B.html
|
|
|